241:
the neurons type and location. This can manifest in different ways. The simplest way is whether the cortical neuron responds only to the deflection of one whisker, or to the deflection of many whiskers. Neurons in layer 4 barrels tend to strongly or exclusively respond to one whisker, while neurons in other layers are less strongly tuned and can respond to multiple whiskers. Neurons that respond to the deflection of multiple whiskers typically have a primary whisker, to which they respond the most. The difference in response magnitude between deflection of the primary whisker and secondary whiskers can also vary between neurons. Stimulation of multiple whiskers may produce a response that is equal to the sum of the responses if each whisker was stimulated independently, or it may be different. Some neurons show greater responses when multiple neurons are stimulated in sequence, and the sequence may be direction specific.
245:
will fire when the whisker that barrel represents is moved at a rate that is roughly proportional to the angular displacement of the neuron. These neurons also show directional sensitivity; certain neurons will only fire when the whisker is moved in a specific direction. Deflection-based firing neurons can sustain their response throughout the deflection of the whisker. Other neurons respond to the initial deflection, but then quickly return to their previous level of activity. Much of this activity is also modulated by the behaviour of the animal - rats and mice actively move their whiskers to explore their environment, and the response of a neuron to a particular stimulus can vary depending on what the animal is doing.
288:-like processes also seem to be involved. This can be inferred by using transgenic mice where there are changes in the expression of enzymes related to LTP and LTD e.g. calmodulin-dependent protein kinase II (CaMKII) or cyclic-AMP response element binding protein (CREB). In these mice, plasticity is compromised Spike timing rather than frequency may be an important factor. Associative LTP has been demonstrated at layer 4 to layer 2/3 synapses when the layer 4 neuron fires 0-15 ms before the layer 2/3 neuron, and LTD is observed when this timing order is reversed. Such mechanisms could act rapidly to produce plastic changes within hours or days.
166:(whiskers) on the mystacial pad (region where whiskers grow from) of certain mammals, they hypothesized that the barrels were the "cortical correlates of the mystacial vibrissae" and that "one barrel represents one vibrissa". Whereas small non-whisker areas of barrel cortex correspond to large and sometimes overlapping areas of the body, each much larger whisker barrel corresponds to a single whisker. As a result, the whisker barrels are the focus of the majority of barrel cortex research, and 'barrel cortex' is often used to refer primarily to the whisker barrels. Consequently, much of this article focuses on rodent whisker barrel cortex.
187:. In the case of the barrel field, the map is somatotopic - based on the arrangement of body parts. Areas corresponding to the nose and mouth are more rostral and lateral in the map, the forelimb, hindlimb and trunk are more medial, with the forelimb rostral of the hindlimb, and the whisker barrel subfields - the posteromedial barrel subfield, which corresponds to the major facial whiskers (the mystacial vibrissae), and the anteriolateral barrel subfield, which corresponds to the smaller whiskers of the face - are caudal and lateral. Although the whiskers make up a relatively small portion of the animal, they dominate the somatotopic map.
300:
trimming. Dendritic branching is important during prenatal and neonatal development, is involved in plasticity induced by lesions, but is not involved in experience-dependent plasticity. In vivo two-photon microscopy reveals that dendritic spines in mouse barrel cortex are highly dynamic and subject to continuous turnover, and may be associated with formation or deletion of synapses. It is likely that spine turnover is necessary but not sufficient to produce experience-dependent plasticity, and other mechanisms such as axonal remodelling are also needed to explain features such as savings from prior experience.
196:
are organized into 5 rows of 4-7 large whiskers that run close to parallel with the bridge of the nose. The organisation of the mystacial vibrissae and corresponding barrels is so consistent that there is a naming convention to identify each whisker in rats and mice. Rows are designated A to E from top to bottom, and columns of whiskers within each row are numbered from back to front. The first four rows also have an additional whisker behind column 1, which is designated with a lower case letter or a Greek letter (α, β, γ, or δ). These four whiskers are also called straddlers.
228:
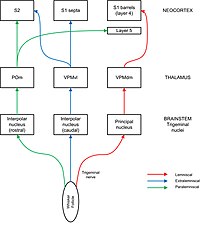
paralemniscal. In the lemniscal pathway, axons from the principal trigeminal nucleus cross over the midline and project to “barreloids” in the thalamus, specifically in the dorsomedial section of the ventroposterior medial nucleus (VPMdm). Neurons in VPMdm project mainly to barrels in layer 4 of primary somatosensory cortex (S1). In the extralemniscal pathway, neurons of the interpolar nucleus project to the ventrolateral section of the ventroposterior medial nucleus (VPMvl). Neurons in VPMvl project to septa between the barrels and to
154:, and each barrel corresponds to a region of the body. Due to this distinctive cellular structure, organisation, and functional significance, the barrel cortex is a useful tool to understand cortical processing and has played an important role in neuroscience. The majority of what is known about corticothalamic processing comes from studying the barrel cortex, and researchers have intensively studied the barrel cortex as a model of
259:
trigeminal nerve) or by ablating, plucking, or trimming some of the facial whiskers. The anatomical structure of the barrels is only affected by lesioning elements of the pathway, but innocuous forms of deprivation can induce rapid changes in the cortical map into adulthood, without any corresponding changes in the barrel structures. Because of their different effects, it seems these two paradigms work by different mechanisms.
232:(S2). The paralemniscal pathway runs from the interpolar trigeminal nucleus via posterior nucleus (POm) of the thalamus to S2 and to diffuse targets in barrel cortex especially layer 5. Each pathway also has secondary projections to other layers within barrel cortex and other regions of cortex, including motor cortex. These different pathways are thought to transmit different modalities of sensory information from the whisker.
118:
175:
205:
25:
270:
different time courses, with the weakening of deprived response preceding the strengthening of spared response, implying that they have different underlying mechanisms. These two effects combine to produce an expansion of the cortical representation of spared whiskers into the representation of adjacent deprived whiskers.
266:. Plucking whiskers in neonatal rats causes a long-lasting expansion of the representation of the spared whisker in layer 4. However, layer 4 plasticity rapidly diminishes if sensory deprivation begins after day 4 of life (P4) whereas representations in layer 2/3 remain highly plastic into adulthood.
240:
The whisker barrel cortex contains different types of neurons that receive input from a range of sources that themselves receive and process an array of different types of information. As a result, neurons of the whisker barrel cortex respond to whisker-related input, but in a way that is specific to
161:
The most distinctive aspect of the barrel field are the whisker barrels. These structures were first discovered by
Woolsey and Van der Loos in 1970. Staining in the whisker barrels is more distinct than that in other areas of the somatosensory cortex. Recognizing that the array was similar to that of
253:
Because the barrel cortex has a well-organised structure that relates clearly to the whisker pad, it has been used extensively as a tool to study sensory processing and development, and the phenomenon of experience-dependent plasticity - changes in the activity, connectivity, and structure of neural
269:
Two cortical processes run alongside each other when barrel cortex is deprived of sensory input from some whiskers to produce representational plasticity. In deprived cortex, neuronal responses to spared whiskers are enhanced and responses to deprived whiskers are weakened. These two processes have
195:
The barrels that correspond to the major facial whiskers (mystacial vibrissae) are contained within the posteromedial barrel subfield (PMBSF). The barrels here are the largest and most elliptical in shape and have a striking topographical organization that is identical to that of the whiskers; they
258:
that allows them to alter the vibrissae to which they respond depending on the rodent's history of tactile experience. Experience-dependent plasticity is commonly studied in the barrel cortex by partially depriving it of sensory input, either by lesioning elements of the afferent pathway (e.g. the
244:
As well as combinations of which whiskers have been stimulated, neurons may also respond to specific types of whisker stimulation. The simplest response, seen in neurons within the layer IV barrel cortex, directly code for whisker displacement. That is to say, that the neuron within a given barrel
299:
It seems intuitively likely that structural changes at the level of axons, dendrite branches, and dendrite spines underlie some of the long-term plastic changes in the cortex. Changes in axon structure have been reported in plasticity following lesions and more recently by studies using whisker
277:
Almost immediately, loss of input to a deprived barrel column leads to a loss of inhibitory firing in that column. This unmasks horizontal excitatory connections from adjacent spared columns. This does not explain longer-lasting plastic changes as the unmasking would disappear immediately if the
227:
carries afferent fibres from the follicles into the brainstem where they connect to neurons in four different trigeminal nerve nuclei: principal, interpolar, oral, and caudal. Projections from the trigeminal nuclei to the thalamus are split into pathways designated lemniscal, extralemniscal, and
295:
amplitude and frequency. The net effect of these changes is to increase the proportion of synaptic input which layer 2/3 neurons in deprived barrels receive from spared barrels. These observations suggest that other, more specific, mechanisms besides LTP/LTD are at play in experience-dependent
475:
Hardingham N, Glazewski S, Pakhotin P, Mizuno K, Chapman PF, Giese KP, Fox K. Neocortical long-term potentiation and experience-dependent synaptic plasticity require alpha-calcium/calmodulin-dependent protein kinase II auto-phosphorylation. J Neurosci. 2003 Jun
178:
Pictomicrograph shows the posteromedial barrel subfield in layer IV of the rat somatosensory cortex. Barrels in the PMBSF are particularly large and distinct. The tissue in the image has been stained with cytochrome oxidase and is 50μm
1128:"Alwis DS, Yan EB, Johnstone V, et al. Environmental enrichment attenuates traumatic brain injury: Induced neuronal hyperexcitability in supragranular layers of sensory cortex. J Neurotrauma. 2016;33(11). doi:10.1089/neu.2014.3774"
1414:
Woolsey, TA; Van Der Loos, H (1970). "The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex: The description of a cortical field composed of discrete cytoarchitectonic units".
216:, that is, they are collected into cylindrical shapes that are narrowed at the top and bottom. The centre of the barrel is designated the hollow, and the spaces between the barrels are the septa (singular: septum)
121:
Pictomicrograph shows the barrel field in layer IV of the rat somatosensory cortex. Each barrel receives input from one whisker. The tissue in the image has been stained with cytochrome oxidase and is 50μm
223:
and the thalamus. Barrel like divisions can be seen in some, but not all parts of the trigeminal nuclei (where they are called barrelets) and the thalamus (where they are called barreloids). The
1168:
Burns (2019) Temporal neuronal activity patterns in barrel cortex to simple and complex stimuli and the effects of traumatic brain injury. Monash
University. Thesis. 10.26180/5b7166ad13e47
1011:
Cheetham CE, Hammond MS, MacFarlane R, Finnerty GT (2008) Altered sensory experience induces targeted rewiring of local excitatory connections in mature neocortex. J Neurosci (in press).
273:
It is likely that several different mechanisms are involved in producing experience-dependent plasticity in a whisker deprivation protocol (adapted from
Feldman and Brecht, 2005 ):
308:, where environmental enrichment of stimuli has been shown to induce plasticity/recovery and patterns of temporal coding have been altered via plasticity and recovery mechanisms.
1344:
Hoover, JE, Hoffer, ZS, Alloway, KD (2003). "Projections From
Primary Somatosensory Cortex to the Neostriatum: The Role of Somatotopic Continuity in Corticostriatal Convergence".
1447:
1594:
1077:"Carron SF, Alwis DS, Rajan R. Traumatic Brain Injury and Neuronal Functionality Changes in Sensory Cortex. Front Syst Neurosci. 2016;10(June):47. doi:10.3389/fnsys.2016.00047"
43:
1024:
Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K (2002). "Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex".
441:
Swadlow HA (1991). "Efferent neurons and suspected interneurons in second somatosensory cortex of the awake rabbit: receptive fields and axonal properties".
1278:"Neural cell adhesion molecule, NCAM, regulates thalamocortical axon pathfinding and the organization of the cortical somatosensory representation in mouse"
1383:
Swadlow HA (1989). "Efferent neurons and suspected interneurons in S-1 vibrissa cortex of the awake rabbit: receptive fields and axonal properties".
150:
and are separated from each other by lighter areas called septa. These dark-staining regions are a major target for somatosensory inputs from the
1569:
1332:
1483:
583:
Glazewski S, Fox K (1996). "Time course of experience-dependent synaptic potentiation anddepression in barrel cortex of adolescent rats".
766:
Glazewski S, Chen CM, Silva A, Fox K (1996). "Requirement for alpha-CaMKII in experience dependent plasticity of the barrel cortex".
292:
61:
1579:
1239:
Diamond ME, von
Heimendahl M, Knutsen PM, Kleinfeld D, Ahissar E (2008). "'Where' and 'what' in the whisker sensorimotor system".
1194:
Bosman LW, Houweling AR, Owens CB, Tanke N, Shevchouk OT, Rahmati N, Teunissen WH, Ju C, Gong W, Koekkoek SK, De Zeeuw CI (2011).
100:
1539:
1639:
229:
184:
907:
Finnerty GT, Roberts LS, Connors BW (1999). "Sensory experience modifies the short-term dynamics of neocortical synapses".
1448:"Comparative anatomical studies of the SmL face cortex with special reference to the occurrence of "barrels" in layer IV"
719:"Sensory loss by selected whisker removal produces immediate disinhibition in the somatosensory cortex of behaving rats"
1544:
619:
1126:
Alwis, D. S.; Yan, E. B.; Johnstone, V.; Carron, S.; Hellewell, S.; Morganti-Kossmann, M. C.; Rajan, R. (2016).
1644:
819:"Impaired experiencedependent plasticity in barrel cortex of mice lacking the alpha and delta isoforms of CREB"
220:
305:
281:
1619:
1033:
967:
916:
775:
285:
147:
131:
255:
860:"Timing-based LTP and LTD at vertical inputs to layer II/III pyramidal cells in rat barrel cortex"
486:
Fox K (2002). "Anatomical pathways and molecular mechanisms for plasticity in the barrel cortex".
1475:
1369:
1264:
1057:
991:
940:
889:
799:
699:
653:
511:
155:
1467:
1432:
1400:
1361:
1328:
1309:
1256:
1227:
1147:
1108:
1049:
983:
932:
881:
840:
791:
748:
691:
645:
600:
562:
503:
458:
1459:
1424:
1392:
1353:
1299:
1289:
1248:
1217:
1207:
1172:
1139:
1098:
1088:
1041:
975:
924:
871:
830:
783:
738:
730:
683:
635:
592:
552:
542:
495:
450:
224:
1634:
263:
143:
139:
254:
circuits in response to experience. Neurons in the barrel cortex exhibit the property of
1514:
1037:
971:
920:
779:
291:
Sensory deprivation has been demonstrated to cause changes in synaptic dynamics such as
1304:
1277:
1222:
1195:
1103:
1076:
743:
734:
718:
557:
547:
530:
204:
958:
Chklovskii DB, Mel BW, Svoboda K (2004). "Cortical rewiring and information storage".
876:
859:
640:
623:
499:
117:
1628:
1428:
531:"A critical period for experience-dependent synaptic plasticity in rat barrel cortex"
212:
The barrels of the barrel cortex were named because the densities of cells resembled
1554:
1479:
893:
803:
515:
304:
Plasticity and remodelling of barrel cortex has also been studied in the context of
142:
and contains the barrel field. The 'barrels' of the barrel field are regions within
1500:
1373:
1268:
1196:"Anatomical pathways involved in generating and sensing rhythmic whisker movements"
1061:
995:
944:
703:
657:
174:
1549:
1524:
787:
624:"Rapid development and plasticity of layer 2/3 maps in rat barrel cortex in vivo"
1599:
596:
454:
278:
deprived input was reinstated (for example by allowing the whisker to regrow).
1396:
1294:
1276:
Enriquez-Barreto, L; Palazzetti C; Brennaman LH; Maness PF; Fairén A (2012).
1212:
1093:
835:
818:
1504:
1127:
687:
163:
1550:
Sensory
Systems & Neural Engineering Group (Hartmann Lab), Chicago, USA
1365:
1313:
1260:
1231:
1176:
1151:
1112:
1053:
987:
936:
885:
844:
752:
695:
649:
507:
93:
1609:
1463:
1436:
1404:
1357:
1143:
795:
604:
566:
462:
219:
Sensory information flows from whisker follicles to barrel cortex via the
1471:
1166:
151:
87:
1045:
979:
106:
1584:
674:
Feldman DE, Brecht M (2005). "Map plasticity in somatosensory cortex".
208:
Sensory information flows in parallel pathways from whiskers to cortex.
1519:
213:
135:
1530:
Tactile
Perception & Learning Lab (Diamond Lab), Triestes, Italy
1252:
817:
Glazewski S, Barth AL, Wallace H, McKenna M, Silva A, Fox K (1999).
16:
Region of the somatosensory cortex in some rodents and other species
183:
The barrel field, like many regions of cortex, is organised into a
1589:
928:
1564:
1590:
Lab of
Sensory Processing (Petersen Lab), Lausanne, Switzerland
1574:
1540:
146:
that are visibly darker when stained to reveal the presence of
1604:
1534:
18:
1559:
1501:
Stained brain slice images which include the "Barrel cortex"
1529:
1075:
Carron, Simone F.; Alwis, Dasuni S.; Rajan, Ramesh (2016).
262:
Some forms of plasticity in the barrel cortex display a
39:
717:Kelly MK, Carvell GE, Kodger JM, Simons DJ (1999).
86:
81:
76:
34:
may be too technical for most readers to understand
1007:
1005:
1600:Barrel Group (Staiger Lab), Göttingen, Germany
578:
576:
1327:. Cambridge, UK: Cambridge University Press.
8:
381:
379:
334:
332:
1510:Research groups working on barrel cortex:
1446:Woolsey TA, Welker C, Schwartz RH (1975).
669:
667:
1303:
1293:
1221:
1211:
1102:
1092:
1019:
1017:
875:
834:
742:
639:
556:
546:
62:Learn how and when to remove this message
46:, without removing the technical details.
371:
203:
173:
134:that is identifiable in some species of
116:
407:
317:
200:Anatomy and connectivity of the barrels
1545:Barrel cortex group (Fox Lab), Cardiff
396:
104:
73:
44:make it understandable to non-experts
7:
1452:The Journal of Comparative Neurology
191:Barrels of the major facial whiskers
1282:Frontiers in Molecular Neuroscience
429:
385:
349:
324:
735:10.1523/JNEUROSCI.19-20-09117.1999
548:10.1523/JNEUROSCI.12-05-01826.1992
418:
360:
138:and species of at least two other
14:
1555:Helmchen Lab, Zürich, Switzerland
1081:Frontiers in Systems Neuroscience
170:Organisation of the barrel fields
386:Woolsey & Van der Loos, 1970
350:Woolsey & Van der Loos, 1970
101:Anatomical terms of neuroanatomy
23:
338:
249:Experience-dependent plasticity
1595:Schwarz Lab, Tübingen, Germany
236:Whisker barrel neurophysiology
230:secondary somatosensory cortex
1:
1580:Oberländer Lab, Bonn, Germany
1565:Kleinfeld Lab, San Diego, USA
1171:(thesis). Monash University.
1165:THOMAS FRANCIS BURNS (2019).
877:10.1016/s0896-6273(00)00008-8
641:10.1016/s0896-6273(01)00360-9
500:10.1016/s0306-4522(02)00027-1
372:Enriquez-Barreto et al., 2012
1585:O'Connor Lab, Baltimore, USA
1429:10.1016/0006-8993(70)90079-x
788:10.1126/science.272.5260.421
1560:Hires Lab, Los Angeles, USA
1661:
1575:Moore Lab, Providence, USA
1535:Feldman Lab, Berkeley, USA
1520:Barth Lab, Pittsburgh, USA
1346:Journal of Neurophysiology
1610:Svoboda Lab, Seattle, USA
1605:Stanley Lab, Atlanta, USA
597:10.1152/jn.1996.75.4.1714
455:10.1152/jn.1991.66.4.1392
99:
1570:Maravall Lab, Sussex, UK
1397:10.1152/jn.1989.62.1.288
1295:10.3389/fnmol.2012.00076
1213:10.3389/fnint.2011.00053
1094:10.3389/fnsys.2016.00047
1615:Books on barrel cortex
1200:Front. Integr. Neurosci
688:10.1126/science.1115807
221:trigeminal nerve nuclei
1177:10.26180/5b7166ad13e47
1132:Journal of Neurotrauma
836:10.1093/cercor/9.3.249
618:Stern EA, Maravall M,
306:traumatic brain injury
209:
180:
123:
1640:Animal nervous system
1525:Bruno Lab, Oxford, UK
1464:10.1002/cne.901640107
1358:10.1152/jn.01009.2002
1144:10.1089/neu.2014.3774
207:
177:
120:
408:Diamond et al., 2008
325:Woolsey et al., 1975
148:cytochrome c oxidase
132:somatosensory cortex
1515:Ahissar Lab, Israel
1046:10.1038/nature01273
1038:2002Natur.420..788T
980:10.1038/nature03012
972:2004Natur.431..782C
921:1999Natur.400..367F
858:Feldman DE (2000).
780:1996Sci...272..421G
419:Bosman et al., 2011
397:Bosman et al., 2011
361:Hoover et al., 2003
256:synaptic plasticity
130:is a region of the
1620:Barrel Cortex book
210:
181:
156:neocortical column
124:
1505:BrainMaps project
1334:978-0-521-85217-3
1138:(11): 1084–1101.
1032:(6917): 788–794.
966:(7010): 782–788.
915:(6742): 367–371.
774:(5260): 421–423.
682:(5749): 810–815.
476:1;23(11):4428-36.
144:cortical layer IV
115:
114:
110:
72:
71:
64:
1652:
1490:
1488:
1482:. Archived from
1440:
1408:
1377:
1352:(3): 1576–1587.
1338:
1317:
1307:
1297:
1272:
1241:Nat Rev Neurosci
1235:
1225:
1215:
1181:
1180:
1162:
1156:
1155:
1123:
1117:
1116:
1106:
1096:
1072:
1066:
1065:
1021:
1012:
1009:
1000:
999:
955:
949:
948:
904:
898:
897:
879:
855:
849:
848:
838:
814:
808:
807:
763:
757:
756:
746:
714:
708:
707:
671:
662:
661:
643:
615:
609:
608:
591:(4): 1714–1729.
580:
571:
570:
560:
550:
541:(5): 1826–1838.
526:
520:
519:
483:
477:
473:
467:
466:
449:(4): 1392–1409.
438:
432:
427:
421:
416:
410:
405:
399:
394:
388:
383:
374:
369:
363:
358:
352:
347:
341:
336:
327:
322:
225:trigeminal nerve
107:edit on Wikidata
74:
67:
60:
56:
53:
47:
27:
26:
19:
1660:
1659:
1655:
1654:
1653:
1651:
1650:
1649:
1645:Neuroplasticity
1625:
1624:
1497:
1486:
1445:
1413:
1382:
1343:
1335:
1323:Fox, K (2008).
1322:
1275:
1253:10.1038/nrn2411
1238:
1193:
1190:
1185:
1184:
1164:
1163:
1159:
1125:
1124:
1120:
1074:
1073:
1069:
1023:
1022:
1015:
1010:
1003:
957:
956:
952:
906:
905:
901:
857:
856:
852:
816:
815:
811:
765:
764:
760:
729:(20): 9117–25.
716:
715:
711:
673:
672:
665:
617:
616:
612:
582:
581:
574:
528:
527:
523:
485:
484:
480:
474:
470:
440:
439:
435:
428:
424:
417:
413:
406:
402:
395:
391:
384:
377:
370:
366:
359:
355:
348:
344:
337:
330:
323:
319:
314:
264:critical period
251:
238:
202:
193:
185:topographic map
172:
111:
68:
57:
51:
48:
40:help improve it
37:
28:
24:
17:
12:
11:
5:
1658:
1656:
1648:
1647:
1642:
1637:
1627:
1626:
1623:
1622:
1613:
1612:
1607:
1602:
1597:
1592:
1587:
1582:
1577:
1572:
1567:
1562:
1557:
1552:
1547:
1542:
1537:
1532:
1527:
1522:
1517:
1508:
1507:
1496:
1495:External links
1493:
1492:
1491:
1489:on 2012-12-23.
1442:
1441:
1423:(2): 205–242.
1417:Brain Research
1410:
1409:
1391:(1): 288–308.
1385:J Neurophysiol
1379:
1378:
1340:
1339:
1333:
1319:
1318:
1273:
1247:(8): 601–612.
1236:
1189:
1186:
1183:
1182:
1157:
1118:
1067:
1013:
1001:
950:
899:
850:
829:(3): 249–256.
809:
758:
709:
663:
634:(2): 305–315.
610:
585:J Neurophysiol
572:
529:Fox K (1992).
521:
494:(4): 799–814.
478:
468:
443:J Neurophysiol
433:
422:
411:
400:
389:
375:
364:
353:
342:
328:
316:
315:
313:
310:
302:
301:
297:
289:
279:
250:
247:
237:
234:
201:
198:
192:
189:
171:
168:
113:
112:
103:
97:
96:
91:
84:
83:
79:
78:
70:
69:
31:
29:
22:
15:
13:
10:
9:
6:
4:
3:
2:
1657:
1646:
1643:
1641:
1638:
1636:
1633:
1632:
1630:
1621:
1618:
1617:
1616:
1611:
1608:
1606:
1603:
1601:
1598:
1596:
1593:
1591:
1588:
1586:
1583:
1581:
1578:
1576:
1573:
1571:
1568:
1566:
1563:
1561:
1558:
1556:
1553:
1551:
1548:
1546:
1543:
1541:
1538:
1536:
1533:
1531:
1528:
1526:
1523:
1521:
1518:
1516:
1513:
1512:
1511:
1506:
1502:
1499:
1498:
1494:
1485:
1481:
1477:
1473:
1469:
1465:
1461:
1457:
1453:
1449:
1444:
1443:
1438:
1434:
1430:
1426:
1422:
1418:
1412:
1411:
1406:
1402:
1398:
1394:
1390:
1386:
1381:
1380:
1375:
1371:
1367:
1363:
1359:
1355:
1351:
1347:
1342:
1341:
1336:
1330:
1326:
1325:Barrel Cortex
1321:
1320:
1315:
1311:
1306:
1301:
1296:
1291:
1287:
1283:
1279:
1274:
1270:
1266:
1262:
1258:
1254:
1250:
1246:
1242:
1237:
1233:
1229:
1224:
1219:
1214:
1209:
1205:
1201:
1197:
1192:
1191:
1187:
1178:
1174:
1170:
1169:
1161:
1158:
1153:
1149:
1145:
1141:
1137:
1133:
1129:
1122:
1119:
1114:
1110:
1105:
1100:
1095:
1090:
1086:
1082:
1078:
1071:
1068:
1063:
1059:
1055:
1051:
1047:
1043:
1039:
1035:
1031:
1027:
1020:
1018:
1014:
1008:
1006:
1002:
997:
993:
989:
985:
981:
977:
973:
969:
965:
961:
954:
951:
946:
942:
938:
934:
930:
929:10.1038/22553
926:
922:
918:
914:
910:
903:
900:
895:
891:
887:
883:
878:
873:
869:
865:
861:
854:
851:
846:
842:
837:
832:
828:
824:
820:
813:
810:
805:
801:
797:
793:
789:
785:
781:
777:
773:
769:
762:
759:
754:
750:
745:
740:
736:
732:
728:
724:
720:
713:
710:
705:
701:
697:
693:
689:
685:
681:
677:
670:
668:
664:
659:
655:
651:
647:
642:
637:
633:
629:
625:
621:
614:
611:
606:
602:
598:
594:
590:
586:
579:
577:
573:
568:
564:
559:
554:
549:
544:
540:
536:
532:
525:
522:
517:
513:
509:
505:
501:
497:
493:
489:
482:
479:
472:
469:
464:
460:
456:
452:
448:
444:
437:
434:
431:
430:Swadlow, 1989
426:
423:
420:
415:
412:
409:
404:
401:
398:
393:
390:
387:
382:
380:
376:
373:
368:
365:
362:
357:
354:
351:
346:
343:
340:
335:
333:
329:
326:
321:
318:
311:
309:
307:
298:
294:
290:
287:
283:
280:
276:
275:
274:
271:
267:
265:
260:
257:
248:
246:
242:
235:
233:
231:
226:
222:
217:
215:
206:
199:
197:
190:
188:
186:
176:
169:
167:
165:
159:
157:
153:
149:
145:
141:
137:
133:
129:
128:barrel cortex
119:
108:
102:
98:
95:
92:
89:
85:
80:
77:Barrel cortex
75:
66:
63:
55:
45:
41:
35:
32:This article
30:
21:
20:
1614:
1509:
1484:the original
1458:(1): 79–94.
1455:
1451:
1420:
1416:
1388:
1384:
1349:
1345:
1324:
1285:
1281:
1244:
1240:
1203:
1199:
1167:
1160:
1135:
1131:
1121:
1084:
1080:
1070:
1029:
1025:
963:
959:
953:
912:
908:
902:
870:(1): 45–56.
867:
863:
853:
826:
823:Cereb Cortex
822:
812:
771:
767:
761:
726:
722:
712:
679:
675:
631:
627:
613:
588:
584:
538:
534:
524:
491:
488:Neuroscience
487:
481:
471:
446:
442:
436:
425:
414:
403:
392:
367:
356:
345:
320:
303:
272:
268:
261:
252:
243:
239:
218:
211:
194:
182:
160:
127:
125:
58:
52:January 2016
49:
33:
723:J. Neurosci
296:plasticity.
82:Identifiers
1629:Categories
1188:References
535:J Neurosci
620:Svoboda K
339:Fox, 2008
164:vibrissae
1480:12374398
1366:12611938
1314:22723769
1261:18641667
1232:22065951
1152:26715144
1113:27313514
1054:12490942
988:15483599
937:10432115
894:17650728
886:10939330
845:10355905
804:84433995
753:10516329
696:16272113
650:11502260
622:(2001).
516:39423181
508:12031405
152:thalamus
88:NeuroLex
1503:at the
1437:4904874
1405:2754479
1374:3002038
1305:3378950
1269:6450408
1223:3207327
1104:4889613
1062:4341820
1034:Bibcode
996:4430167
968:Bibcode
945:4413560
917:Bibcode
796:8602534
776:Bibcode
768:Science
744:6782760
704:2892382
676:Science
658:2819415
605:8727408
567:1578273
558:6575898
463:1761989
214:barrels
136:rodents
38:Please
1635:Memory
1478:
1472:809494
1470:
1435:
1403:
1372:
1364:
1331:
1312:
1302:
1288:: 76.
1267:
1259:
1230:
1220:
1206:: 53.
1150:
1111:
1101:
1087:: 47.
1060:
1052:
1026:Nature
994:
986:
960:Nature
943:
935:
909:Nature
892:
884:
864:Neuron
843:
802:
794:
751:
741:
702:
694:
656:
648:
628:Neuron
603:
565:
555:
514:
506:
461:
284:- and
179:thick.
140:orders
122:thick.
94:nlx_81
1487:(pdf)
1476:S2CID
1370:S2CID
1265:S2CID
1058:S2CID
992:S2CID
941:S2CID
890:S2CID
800:S2CID
700:S2CID
654:S2CID
512:S2CID
312:Notes
105:[
1468:PMID
1433:PMID
1401:PMID
1362:PMID
1329:ISBN
1310:PMID
1257:PMID
1228:PMID
1148:PMID
1109:PMID
1050:PMID
984:PMID
933:PMID
882:PMID
841:PMID
792:PMID
749:PMID
692:PMID
646:PMID
601:PMID
563:PMID
504:PMID
459:PMID
293:EPSP
162:the
126:The
1460:doi
1456:164
1425:doi
1393:doi
1354:doi
1300:PMC
1290:doi
1249:doi
1218:PMC
1208:doi
1173:doi
1140:doi
1099:PMC
1089:doi
1042:doi
1030:420
976:doi
964:431
925:doi
913:400
872:doi
831:doi
784:doi
772:272
739:PMC
731:doi
684:doi
680:310
636:doi
593:doi
553:PMC
543:doi
496:doi
492:111
451:doi
286:LTD
282:LTP
42:to
1631::
1474:.
1466:.
1454:.
1450:.
1431:.
1421:17
1419:.
1399:.
1389:62
1387:.
1368:.
1360:.
1350:89
1348:.
1308:.
1298:.
1284:.
1280:.
1263:.
1255:.
1243:.
1226:.
1216:.
1202:.
1198:.
1146:.
1136:33
1134:.
1130:.
1107:.
1097:.
1085:10
1083:.
1079:.
1056:.
1048:.
1040:.
1028:.
1016:^
1004:^
990:.
982:.
974:.
962:.
939:.
931:.
923:.
911:.
888:.
880:.
868:27
866:.
862:.
839:.
825:.
821:.
798:.
790:.
782:.
770:.
747:.
737:.
727:19
725:.
721:.
698:.
690:.
678:.
666:^
652:.
644:.
632:31
630:.
626:.
599:.
589:75
587:.
575:^
561:.
551:.
539:12
537:.
533:.
510:.
502:.
490:.
457:.
447:66
445:.
378:^
331:^
158:.
90:ID
1462::
1439:.
1427::
1407:.
1395::
1376:.
1356::
1337:.
1316:.
1292::
1286:5
1271:.
1251::
1245:9
1234:.
1210::
1204:5
1179:.
1175::
1154:.
1142::
1115:.
1091::
1064:.
1044::
1036::
998:.
978::
970::
947:.
927::
919::
896:.
874::
847:.
833::
827:9
806:.
786::
778::
755:.
733::
706:.
686::
660:.
638::
607:.
595::
569:.
545::
518:.
498::
465:.
453::
109:]
65:)
59:(
54:)
50:(
36:.
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.