219:
functionally categories into channel or transporter. They share homodimeric structure with independent ion permeation pathway in each of the subunit. Based on functional characterization, there are two known gating mechanism: protopore and common gating. The protopore gating, also known as fast gating, is associated with occlusion of the pore via side-chain of conserved glutamate. While the common gating, also known as the slow gating, inactivated or reactivates both pores through unknown mechanism. This family either transports two chloride for one proton or simply allows flux down its electrochemical gradient. With this channel the correct depolarization and repolarization via chloride ions is essential for propagation of an action potential.
210:
gradient. This influx of calcium subsequently is what causes the neurotransmitter vesicles to fuse with the presynaptic membrane. The calcium ions initiate the interaction of obligatory cofactor proteins with SNARE proteins to form a SNARE complex. These SNARE complexes mediate vesicle fusion by pulling the membranes together, leaking the neurotransmitters into the synaptic cleft. The neurotransmitter molecules can then signal the next cell via receptors on the post synaptic membrane. These receptors can either act as ion channels or GPCR (G-Protein
Coupled Receptors). In general the neurotransmitter can either cause an excitatory or inhibitory response, depending on what occurs at the receptor.
309:. Recent studies have suggested a molecular dynamics simulation-based method to determine gating charge by measuring electrical capacitor properties of membrane-embedded proteins. Activity of ion channels located in the plasma membrane can be measured by simply attaching a glass capillary electrode continuously with the membrane. Other ion channels located in the membranes of mitochondria, lysosomes, and the Golgi apparatus can be measured by an emergent technique that involves the use of an artificial bilayer lipid membrane attached to a 16 electrode device that measures electrical activity.
201:
282:
257:
142:
main ion channels responsible for action potentials. Being complex, they are made of bigger α subunits that are then paired with two smaller β subunits. They contain transmembrane segments known as S1-6. The charged S4 segments are the channels voltage sensors. When exposed to a certain minimum potential difference, the S4 segments move across the membrane. This causes movement of the S4-S5 linker, which causes the S5-S6 linker to twist and opens the channel.
115:
98:), and 'deactivation' is the opposite process of the activation gate closing in response to the inside of the membrane becoming more negative (repolarization). 'Inactivation' is the closing of the inactivation gate, and occurs in response to the voltage inside the membrane becoming more positive, but more slowly than activation. 'Reactivation' is the opposite of inactivation, and is the process of reopening the inactivation gate.
129:, moving these voltage-sensing domains. This changes the conformation of other elements of the channel to either the open or closed position. When they move from the closed position to the open position, this is called "activation." Voltage-gated ion channels underlie many of the electrical behaviors of the cell, including action potentials, resting membrane potentials, and synaptic transmission.
17:
231:
through the channels down their concentration gradient. Ligand-gated ion channels are responsible for fast synaptic transmission in the nervous system and at the neuromuscular junction. Each ligand-gated ion channel has a wide range of receptors with differing biophysical properties as well as patterns of expression in the nervous system.
209:
Neurotransmitters are initially stored and synthesized in vesicles at the synapse of a neuron. When an action potential occurs in a cell, the electrical signal reaches the presynaptic terminal and the depolarization causes calcium channels to open, releasing calcium to travel down its electrochemical
141:
Voltage Gated Sodium (Na) channels are significant when it comes to propagating the action potentials in neurons and other excitable cells, mostly being used for the propagation of action potential in axons, muscle fibers and the neural somatodendritic compartment. Sodium(Na) channels are some of the
272:
N-terminus. The ball domain is electrostatically attracted to the inner channel domain. When the ion channel is activated, the inner channel domain is exposed, and within milliseconds the chain will fold and the ball will enter the channel, occluding ion permeation. The channel returns to its closed
239:
Inactivation is when the flow of ions is blocked by a mechanism other than the closing of the channel. A channel in its open state may stop allowing ions to flow through, or a channel in its closed state may be preemptively inactivated to prevent the flow of ions. Inactivation typically occurs when
196:
Calcium (Ca) channels regulate the release of neurotransmitters at synapses, control the shape of action potentials made by sodium channels, and in some neurons, generate action potentials. Calcium channels consist of six transmembrane helices. S4 acts as the voltage sensor by rotating when exposed
296:
In voltage gated potassium channels, the reverse is true, and deactivation slows the channel's recovery from activation. The closed conformation is assumed by default, and involves the partial straightening of helix VI by the IV-V linker. The mechanisms that cause opening and closing are not fully
150:
Potassium (K) channels play a large role in setting the resting membrane potential. When the cell membrane depolarizes, the intracellular part of the channel becomes positively charged, which causes the channel's open configuration to become a more stable state than the closed configuration. There
230:
are found on postsynaptic neurons. By default, they assume their closed conformation. When the presynaptic neuron releases neurotransmitters at the end of an action potential, they bind to ligand-gated ion channels. This causes the channels to assume their open conformation, allowing ions to flow
247:
In sodium channels, inactivation appears to be the result of the actions of helices III-VI, with III and IV acting as a sort of hinged lid that block the channel. The exact mechanism is poorly understood, but seems to rely on a particle that has a high affinity for the exposed inside of the open
132:
Voltage-gated ion channels are often specific to ions, including Na, K, Ca, and Cl. Each of these ions plays an important role in the electrical behavior of the cell. The gates also have unique properties with important physiological implications. For example, Na channels open and close rapidly,
218:
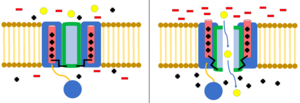
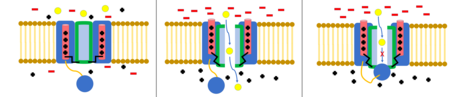
Chloride channels are another group of voltage gated ion channels, of which are less understood. They are involved with processes such as skeletal and cardiac smooth muscle, cell volume regulation, the cell cycle, and apoptosis. One major family of chloride proteins are called CLC proteins,
89:
are often described as having four gating processes: activation, deactivation, inactivation, and reactivation (also called 'recovery from inactivation'). Activation is the process of opening the activation gate, which occurs in response to the voltage inside the cell membrane (the
268:, also known as N-type inactivation or hinged lid inactivation, is a gating mechanism for some voltage-gated ion channels. Voltage-gated ion channels are composed of 4 α subunits, one or more of which will have a ball domain located on its
289:
Deactivation is the return of an ion channel to its closed conformation. For voltage-gated channels this occurs when the voltage differential that originally caused the channel to open returns to its resting value.
204:
Calcium release causes a strong attraction between multiple proteins including synaptobrevin and SNARE proteins to pull the neurotransmitter vesicle to the membrane and release its contents into the synaptic
47:
to ions and do not conduct electrical current. When ion channels are in their open state, they conduct electrical current by allowing specific types of ions to pass through them, and thus, across the
125:
open and close in response to the electrical potential across the cell membrane. Portions of the channel domain act as voltage sensors. As the membrane potential changes, this results in changes in
118:
Voltage-gated ion channel. When the membrane is polarized, the voltage sensing domain of the channel shifts, opening the channel to ion flow (ions represented by yellow circles).
133:
while K gates open and close much more slowly. The difference in speed between these channels underlies the depolarization and repolarization phases of the action potential.
1274:
285:
As the membrane potential returns to its resting value, the voltage differential is not sufficient to keep the channel in its open state, causing the channel to close.
297:
understood. The closed conformation appears to be a higher energy conformation than the open conformation, which may also help explain how the ion channel activates.
260:
Voltage-gated ion channel in its closed, open, and inactivated states. The inactivated channel is still in its open state, but the ball domain blocks ion permeation.
166:
posits that the S3 and S4 helices of the channel form "paddles" that move through the depolarized membrane and pull the S5 helix away from the channel's opening.
792:
Purves, Dale; Augustine, George J.; Fitzpatrick, David; Katz, Lawrence C.; LaMantia, Anthony-Samuel; McNamara, James O.; Williams, S. Mark (2001).
2132:
Kamiya, Koki; Osaki, Toshihisa; Nakao, Kenji; Kawano, Ryuji; Fujii, Satoshi; Misawa, Nobuo; Hayakawa, Masatoshi; Takeuchi, Shoji (2018-11-30).
82:
of gating. Some drugs and many ion channel toxins act as 'gating modifiers' of voltage-gated ion channels by changing the kinetics of gating.
1309:
591:
360:
846:
Mantegazza, Massimo; Catterall, William A. (2012), Noebels, Jeffrey L.; Avoli, Massimo; Rogawski, Michael A.; Olsen, Richard W. (eds.),
58:
A variety of cellular changes can trigger gating, depending on the ion channel, including changes in voltage across the cell membrane (
173:
posits that a focused electric field causes charged particles to move across the channel with only a small movement of the S4 helix.
1781:
Yang, Kefan; Coburger, Ina; Langner, Johanna M.; Peter, Nicole; Hoshi, Toshinori; Schönherr, Roland; Heinemann, Stefan H. (2019).
44:
101:
These voltage-dependent changes in function are critical for a large number of processes in excitable and nonexcitable cells.
248:
channel. Rapid inactivation allows the channel to halt the flow of sodium very shortly after assuming its open conformation.
1895:"Molecular dynamics of the sodium channel pore vary with gating: interactions between P-segment motions and inactivation"
2203:
2134:"Electrophysiological measurement of ion channels on plasma/organelle membranes using an on-chip lipid bilayer system"
265:
180:
posits that the S4 and S5 helices both rotate, and the S4-S5 linker causes the S6 helix to move, opening the channel.
2208:
200:
122:
59:
281:
878:
Sula, Altin; Booker, Jennifer; Ng, Leo C. T.; Naylor, Claire E.; DeCaen, Paul G.; Wallace, B. A. (2017-02-16).
256:
227:
63:
381:
Machtens, Jan-Philipp; Briones, Rodolfo; Alleva, Claudia; de Groot, Bert L.; Fahlke, Christoph (2017-04-11).
1671:
Kuo, Chung-Chin; Bean, Bruce P. (1994-04-01). "Na+ channels must deactivate to recover from inactivation".
1783:"Modulation of K+ channel N-type inactivation by sulfhydration through hydrogen sulfide and polysulfides"
78:). The rate at which any of these gating processes occurs in response to these triggers are known as the
2213:
1268:
306:
2145:
2077:
1614:
1296:, Handbook of Experimental Pharmacology, vol. 283, Berlin, Heidelberg: Springer, pp. 1–34,
1166:
Yoon, Tae-Young; Lu, Xiaobing; Diao, Jiajie; Lee, Soo-Min; Ha, Taekjip; Shin, Yeon-Kyun (June 2008).
891:
536:
469:
394:
126:
343:
Alberts, Bruce; Bray, Dennis; Lewis, Julian; Raff, Martin; Roberts, Keith; Watson, James D. (1994).
1704:
1436:
607:
Waszkielewicz, A.M; Gunia, A; Szkaradek, N; Słoczyńska, K; Krupińska, S; Marona, H (April 2013).
459:
323:
91:
79:
55:. Gating is the process by which an ion channel transitions between its open and closed states.
2179:
2161:
2111:
2093:
2046:
2028:
1989:
1971:
1932:
1914:
1875:
1857:
1812:
1763:
1745:
1696:
1688:
1650:
1632:
1532:
1493:
1475:
1428:
1420:
1379:
1361:
1315:
1305:
1246:
1205:
1187:
1148:
1130:
1088:
1070:
1031:
982:
964:
925:
907:
855:
820:
765:
747:
703:
685:
646:
628:
587:
564:
505:
487:
428:
410:
356:
352:
241:
25:
2169:
2153:
2101:
2085:
2036:
2020:
1979:
1963:
1922:
1906:
1865:
1849:
1802:
1794:
1753:
1735:
1680:
1640:
1622:
1581:
1573:
1540:
1524:
1483:
1467:
1410:
1369:
1351:
1297:
1254:
1238:
1195:
1179:
1138:
1122:
1078:
1062:
1021:
1013:
972:
956:
915:
899:
755:
739:
693:
677:
636:
620:
554:
544:
495:
477:
418:
402:
86:
114:
318:
293:
In voltage-gated sodium channels, deactivation is necessary to recover from inactivation.
71:
66:), changes in temperature, stretching or deformation of the cell membrane, addition of a
2149:
2081:
1618:
895:
540:
473:
398:
2174:
2133:
2106:
2065:
2041:
2024:
2008:
1984:
1951:
1927:
1910:
1894:
1870:
1837:
1807:
1645:
1602:
1586:
1577:
1561:
1545:
1528:
1512:
1488:
1455:
1374:
1339:
1259:
1242:
1226:
1200:
1167:
1143:
1110:
1083:
1050:
1026:
1001:
977:
944:
920:
879:
760:
727:
698:
665:
641:
608:
423:
382:
95:
52:
1758:
1723:
1399:"CLC Chloride Channels and Transporters: Structure, Function, Physiology, and Disease"
500:
447:
2197:
1684:
1017:
664:
Ahern, Christopher A.; Payandeh, Jian; Bosmans, Frank; Chanda, Baron (January 2016).
559:
524:
345:
48:
1893:
Bénitah, J. P.; Chen, Z.; Balser, J. R.; Tomaselli, G. F.; Marbán, E. (1999-03-01).
1708:
1440:
960:
187:
is an average of the above models that helps reconcile them with experimental data.
37:
1967:
1471:
743:
159:
posits that the potassium channel opens due to a screwing motion by its S4 helix.
40:. This change in conformation is a response to changes in transmembrane voltage.
1126:
1066:
624:
2157:
1415:
1398:
812:
793:
452:
Proceedings of the
National Academy of Sciences of the United States of America
1798:
945:"Structure and function of voltage-gated sodium channels at atomic resolution"
847:
406:
33:
2165:
2097:
2032:
1975:
1918:
1861:
1749:
1692:
1636:
1536:
1479:
1424:
1365:
1250:
1191:
1134:
1074:
968:
911:
751:
689:
632:
491:
414:
20:
An animated representation of the molecular structure of a simple ion channel
1740:
1627:
549:
269:
75:
67:
2183:
2115:
1993:
1936:
1816:
1767:
1654:
1497:
1432:
1383:
1319:
1209:
1152:
1092:
1035:
986:
929:
859:
824:
769:
707:
650:
568:
509:
482:
432:
383:"Gating Charge Calculations by Computational Electrophysiology Simulations"
2066:"The pore of voltage-gated potassium ion channels is strained when closed"
2050:
1879:
1853:
1700:
681:
16:
1301:
1356:
1338:
Park, Eunyong; MacKinnon, Roderick (2018-05-29). Csanády, László (ed.).
903:
464:
2089:
2009:"Deactivation Retards Recovery from Inactivation in Shaker K+ Channels"
1952:"Mechanisms of closed-state inactivation in voltage-gated ion channels"
1456:"CLC channels and transporters: proteins with borderline personalities"
1289:
728:"Mechanisms of closed-state inactivation in voltage-gated ion channels"
448:"Ion channel gating: A first-passage time analysis of the Kramers type"
1183:
848:"Voltage-Gated Na+ Channels: Structure, Function, and Pathophysiology"
43:
When ion channels are in a 'closed' (non-conducting) state, they are
854:(4th ed.), National Center for Biotechnology Information (US),
1838:"N-type inactivation and the S4-S5 region of the Shaker K+ channel"
1290:"Biophysical and Pharmacological Insights to CLC Chloride Channels"
666:"The hitchhiker's guide to the voltage-gated sodium channel galaxy"
609:"Ion Channels as Drug Targets in Central Nervous System Disorders"
273:
state, blocking the channel domain, and the ball leaves the pore.
255:
199:
113:
94:) becoming more positive with respect to the outside of the cell (
15:
1288:
Kwon, Hwoi Chan; Fairclough, Robert H.; Chen, Tsung-Yu (2022),
1000:
Grizel, A. V.; Glukhov, G. S.; Sokolova, O. S. (Oct–Dec 2014).
1168:"Complexin and Ca 2+ stimulate SNARE-mediated membrane fusion"
1002:"Mechanisms of Activation of Voltage-Gated Potassium Channels"
197:
to certain membrane potentials, thereby opening the channel.
880:"The complete structure of an activated open sodium channel"
523:
Cesare P, Moriondo A, Vellani V, McNaughton PA (July 1999).
1340:"Structure of the CLC-1 chloride channel from Homo sapiens"
74:), and interaction with other molecules in the cell (e.g.,
1836:
Holmgren, M.; Jurman, M. E.; Yellen, G. (September 1996).
1782:
1511:
Alexander, SPH; Mathie, A; Peters, JA (November 2011).
1724:"Overview of the voltage-gated sodium channel family"
1603:"Na channel inactivation from open and closed states"
1454:
Accardi, Alessio; Picollo, Alessandra (August 2010).
811:
Grider, Michael H.; Glaubensklee, Carolyn S. (2019),
2064:
Fowler, Philip W.; Sansom, Mark S. P. (2013-05-21).
1950:Bähring, Robert; Covarrubias, Manuel (2011-01-28).
726:Bähring, Robert; Covarrubias, Manuel (2011-02-01).
1460:Biochimica et Biophysica Acta (BBA) - Biomembranes
344:
151:are a few models of potassium channel activation:
36:) or closing (by deactivation or inactivation) of
1397:Jentsch, Thomas J.; Pusch, Michael (2018-07-01).
1787:Pflügers Archiv - European Journal of Physiology
240:the cell membrane depolarize, and ends when the
1607:Proceedings of the National Academy of Sciences
62:), chemicals interacting with the ion channel (
1560:Alexander, SPH; Mathie, A; Peters, JA (2011).
1666:
1664:
1111:"Calcium Control of Neurotransmitter Release"
8:
1273:: CS1 maint: DOI inactive as of July 2024 (
852:Jasper's Basic Mechanisms of the Epilepsies
446:Goychuk, Igor; Hänggi, Peter (2002-03-19).
305:Gating charge can be calculated by solving
1722:Yu, Frank H; Catterall, William A (2003).
1115:Cold Spring Harbor Perspectives in Biology
1055:Cold Spring Harbor Perspectives in Biology
2173:
2105:
2040:
1983:
1926:
1869:
1806:
1757:
1739:
1644:
1626:
1585:
1544:
1487:
1414:
1373:
1355:
1258:
1199:
1172:Nature Structural & Molecular Biology
1142:
1082:
1025:
976:
919:
759:
697:
640:
558:
548:
499:
481:
463:
422:
280:
335:
1266:
85:The voltage-gated ion channels of the
2127:
2125:
1237:(Suppl 1): S130–S134. November 2009.
1221:
1219:
1104:
1102:
1049:Catterall, William A. (August 2011).
841:
839:
787:
785:
783:
781:
779:
7:
943:Catterall, William A. (2013-11-14).
721:
719:
717:
376:
374:
372:
584:Ion channels of excitable membranes
2025:10.1523/JNEUROSCI.17-10-03436.1997
1911:10.1523/JNEUROSCI.19-05-01577.1999
1578:10.1111/j.1476-5381.2011.01649_4.x
1529:10.1111/j.1476-5381.2011.01649_4.x
1243:10.1111/j.1476-5381.2009.00503_6.x
1109:Südhof, Thomas C. (January 2012).
14:
1842:The Journal of General Physiology
1601:Armstrong, Clay M. (2006-11-21).
670:The Journal of General Physiology
1051:"Voltage-Gated Calcium Channels"
1018:10.32607/20758251-2014-6-4-10-26
1517:British Journal of Pharmacology
1294:Anion Channels and Transporters
1231:British Journal of Pharmacology
178:coordinated movement of helices
2007:Kuo, Chung-Chin (1997-05-15).
961:10.1113/expphysiol.2013.071969
813:"Physiology, Action Potential"
351:. New York: Garland. pp.
1:
586:. Sunderland, Mass: Sinauer.
347:Molecular biology of the cell
1968:10.1113/jphysiol.2010.191965
1685:10.1016/0896-6273(94)90335-2
1472:10.1016/j.bbamem.2010.02.022
794:"Voltage-Gated Ion Channels"
744:10.1113/jphysiol.2010.191965
529:Proc. Natl. Acad. Sci. U.S.A
525:"Ion channels gated by heat"
2013:The Journal of Neuroscience
1899:The Journal of Neuroscience
1562:"Ligand-Gated Ion Channels"
1513:"Ligand-Gated Ion Channels"
1127:10.1101/cshperspect.a011353
1067:10.1101/cshperspect.a003947
625:10.2174/0929867311320100005
613:Current Medicinal Chemistry
252:Ball and chain inactivation
2230:
2158:10.1038/s41598-018-35316-4
1416:10.1152/physrev.00047.2017
123:Voltage-gated ion channels
110:Voltage-gated ion channels
70:group to the ion channel (
60:voltage-gated ion channels
1956:The Journal of Physiology
1799:10.1007/s00424-018-2233-x
819:, StatPearls Publishing,
798:Neuroscience. 2nd Edition
732:The Journal of Physiology
407:10.1016/j.bpj.2017.02.016
228:Ligand-gated ion channels
223:Ligand-gated ion channels
64:ligand-gated ion channels
1741:10.1186/gb-2003-4-3-207
1628:10.1073/pnas.0607603103
1245:(inactive 2024-07-16).
949:Experimental Physiology
550:10.1073/pnas.96.14.7658
32:refers to the opening (
1572:(Suppl 1): S115–S135.
1523:(Suppl 1): S115–S135.
582:Hille, Bertil (2001).
483:10.1073/pnas.052015699
286:
261:
206:
119:
21:
2070:Nature Communications
1854:10.1085/jgp.108.3.195
1403:Physiological Reviews
884:Nature Communications
682:10.1085/jgp.201511492
284:
259:
203:
117:
19:
1302:10.1007/164_2022_594
266:ball and chain model
127:electrostatic forces
2150:2018NatSR...817498K
2082:2013NatCo...4.1872F
1619:2006PNAS..10317991A
1613:(47): 17991–17996.
1357:10.7554/eLife.36629
1227:"Chloride channels"
904:10.1038/ncomms14205
896:2017NatCo...814205S
541:1999PNAS...96.7658C
474:2002PNAS...99.3552G
399:2017BpJ...112.1396M
387:Biophysical Journal
324:Synaptic potentials
157:sliding helix model
2204:Cell communication
2138:Scientific Reports
2090:10.1038/ncomms2858
307:Poisson's equation
287:
262:
207:
120:
92:membrane potential
22:
2209:Electrophysiology
2019:(10): 3436–3444.
1311:978-3-031-51345-9
1184:10.1038/nsmb.1446
738:(Pt 3): 461–479.
619:(10): 1241–1285.
593:978-0-87893-321-1
362:978-0-8153-1620-6
242:resting potential
26:electrophysiology
2221:
2188:
2187:
2177:
2129:
2120:
2119:
2109:
2061:
2055:
2054:
2044:
2004:
1998:
1997:
1987:
1947:
1941:
1940:
1930:
1905:(5): 1577–1585.
1890:
1884:
1883:
1873:
1833:
1827:
1826:
1824:
1823:
1810:
1778:
1772:
1771:
1761:
1743:
1719:
1713:
1712:
1668:
1659:
1658:
1648:
1630:
1598:
1592:
1591:
1589:
1557:
1551:
1550:
1548:
1508:
1502:
1501:
1491:
1466:(8): 1457–1464.
1451:
1445:
1444:
1418:
1409:(3): 1493–1590.
1394:
1388:
1387:
1377:
1359:
1335:
1329:
1328:
1327:
1326:
1285:
1279:
1278:
1272:
1264:
1262:
1223:
1214:
1213:
1203:
1163:
1157:
1156:
1146:
1106:
1097:
1096:
1086:
1046:
1040:
1039:
1029:
997:
991:
990:
980:
940:
934:
933:
923:
875:
869:
868:
867:
866:
843:
834:
833:
832:
831:
808:
802:
801:
789:
774:
773:
763:
723:
712:
711:
701:
661:
655:
654:
644:
604:
598:
597:
579:
573:
572:
562:
552:
520:
514:
513:
503:
485:
467:
458:(6): 3552–3556.
443:
437:
436:
426:
393:(7): 1396–1405.
378:
367:
366:
350:
340:
87:action potential
2229:
2228:
2224:
2223:
2222:
2220:
2219:
2218:
2194:
2193:
2192:
2191:
2131:
2130:
2123:
2063:
2062:
2058:
2006:
2005:
2001:
1949:
1948:
1944:
1892:
1891:
1887:
1835:
1834:
1830:
1821:
1819:
1780:
1779:
1775:
1721:
1720:
1716:
1670:
1669:
1662:
1600:
1599:
1595:
1559:
1558:
1554:
1510:
1509:
1505:
1453:
1452:
1448:
1396:
1395:
1391:
1337:
1336:
1332:
1324:
1322:
1312:
1287:
1286:
1282:
1265:
1225:
1224:
1217:
1165:
1164:
1160:
1108:
1107:
1100:
1048:
1047:
1043:
999:
998:
994:
942:
941:
937:
877:
876:
872:
864:
862:
845:
844:
837:
829:
827:
810:
809:
805:
791:
790:
777:
725:
724:
715:
663:
662:
658:
606:
605:
601:
594:
581:
580:
576:
535:(14): 7658–63.
522:
521:
517:
465:physics/0111187
445:
444:
440:
380:
379:
370:
363:
342:
341:
337:
332:
319:Synaptic gating
315:
303:
279:
254:
237:
225:
216:
194:
185:consensus model
171:transport model
148:
139:
112:
107:
72:phosphorylation
49:plasma membrane
12:
11:
5:
2227:
2225:
2217:
2216:
2211:
2206:
2196:
2195:
2190:
2189:
2121:
2056:
1999:
1962:(3): 461–479.
1942:
1885:
1848:(3): 195–206.
1828:
1793:(4): 557–571.
1773:
1728:Genome Biology
1714:
1679:(4): 819–829.
1660:
1593:
1566:Br J Pharmacol
1552:
1503:
1446:
1389:
1330:
1310:
1280:
1215:
1178:(7): 707–713.
1158:
1121:(1): a011353.
1098:
1061:(8): a003947.
1041:
992:
935:
870:
835:
803:
775:
713:
656:
599:
592:
574:
515:
438:
368:
361:
334:
333:
331:
328:
327:
326:
321:
314:
311:
302:
301:Quantification
299:
278:
275:
253:
250:
236:
233:
224:
221:
215:
212:
193:
190:
189:
188:
181:
174:
167:
160:
147:
144:
138:
135:
111:
108:
106:
103:
96:depolarization
13:
10:
9:
6:
4:
3:
2:
2226:
2215:
2212:
2210:
2207:
2205:
2202:
2201:
2199:
2185:
2181:
2176:
2171:
2167:
2163:
2159:
2155:
2151:
2147:
2143:
2139:
2135:
2128:
2126:
2122:
2117:
2113:
2108:
2103:
2099:
2095:
2091:
2087:
2083:
2079:
2075:
2071:
2067:
2060:
2057:
2052:
2048:
2043:
2038:
2034:
2030:
2026:
2022:
2018:
2014:
2010:
2003:
2000:
1995:
1991:
1986:
1981:
1977:
1973:
1969:
1965:
1961:
1957:
1953:
1946:
1943:
1938:
1934:
1929:
1924:
1920:
1916:
1912:
1908:
1904:
1900:
1896:
1889:
1886:
1881:
1877:
1872:
1867:
1863:
1859:
1855:
1851:
1847:
1843:
1839:
1832:
1829:
1818:
1814:
1809:
1804:
1800:
1796:
1792:
1788:
1784:
1777:
1774:
1769:
1765:
1760:
1755:
1751:
1747:
1742:
1737:
1733:
1729:
1725:
1718:
1715:
1710:
1706:
1702:
1698:
1694:
1690:
1686:
1682:
1678:
1674:
1667:
1665:
1661:
1656:
1652:
1647:
1642:
1638:
1634:
1629:
1624:
1620:
1616:
1612:
1608:
1604:
1597:
1594:
1588:
1583:
1579:
1575:
1571:
1567:
1563:
1556:
1553:
1547:
1542:
1538:
1534:
1530:
1526:
1522:
1518:
1514:
1507:
1504:
1499:
1495:
1490:
1485:
1481:
1477:
1473:
1469:
1465:
1461:
1457:
1450:
1447:
1442:
1438:
1434:
1430:
1426:
1422:
1417:
1412:
1408:
1404:
1400:
1393:
1390:
1385:
1381:
1376:
1371:
1367:
1363:
1358:
1353:
1349:
1345:
1341:
1334:
1331:
1321:
1317:
1313:
1307:
1303:
1299:
1295:
1291:
1284:
1281:
1276:
1270:
1261:
1256:
1252:
1248:
1244:
1240:
1236:
1232:
1228:
1222:
1220:
1216:
1211:
1207:
1202:
1197:
1193:
1189:
1185:
1181:
1177:
1173:
1169:
1162:
1159:
1154:
1150:
1145:
1140:
1136:
1132:
1128:
1124:
1120:
1116:
1112:
1105:
1103:
1099:
1094:
1090:
1085:
1080:
1076:
1072:
1068:
1064:
1060:
1056:
1052:
1045:
1042:
1037:
1033:
1028:
1023:
1019:
1015:
1011:
1007:
1003:
996:
993:
988:
984:
979:
974:
970:
966:
962:
958:
954:
950:
946:
939:
936:
931:
927:
922:
917:
913:
909:
905:
901:
897:
893:
889:
885:
881:
874:
871:
861:
857:
853:
849:
842:
840:
836:
826:
822:
818:
814:
807:
804:
799:
795:
788:
786:
784:
782:
780:
776:
771:
767:
762:
757:
753:
749:
745:
741:
737:
733:
729:
722:
720:
718:
714:
709:
705:
700:
695:
691:
687:
683:
679:
675:
671:
667:
660:
657:
652:
648:
643:
638:
634:
630:
626:
622:
618:
614:
610:
603:
600:
595:
589:
585:
578:
575:
570:
566:
561:
556:
551:
546:
542:
538:
534:
530:
526:
519:
516:
511:
507:
502:
497:
493:
489:
484:
479:
475:
471:
466:
461:
457:
453:
449:
442:
439:
434:
430:
425:
420:
416:
412:
408:
404:
400:
396:
392:
388:
384:
377:
375:
373:
369:
364:
358:
354:
349:
348:
339:
336:
329:
325:
322:
320:
317:
316:
312:
310:
308:
300:
298:
294:
291:
283:
276:
274:
271:
267:
258:
251:
249:
245:
244:is restored.
243:
234:
232:
229:
222:
220:
213:
211:
202:
198:
191:
186:
182:
179:
176:The model of
175:
172:
168:
165:
161:
158:
154:
153:
152:
145:
143:
136:
134:
130:
128:
124:
116:
109:
104:
102:
99:
97:
93:
88:
83:
81:
77:
73:
69:
65:
61:
56:
54:
50:
46:
41:
39:
35:
31:
27:
18:
2214:Ion channels
2144:(1): 17498.
2141:
2137:
2073:
2069:
2059:
2016:
2012:
2002:
1959:
1955:
1945:
1902:
1898:
1888:
1845:
1841:
1831:
1820:. Retrieved
1790:
1786:
1776:
1731:
1727:
1717:
1676:
1672:
1610:
1606:
1596:
1569:
1565:
1555:
1520:
1516:
1506:
1463:
1459:
1449:
1406:
1402:
1392:
1347:
1343:
1333:
1323:, retrieved
1293:
1283:
1269:cite journal
1234:
1230:
1175:
1171:
1161:
1118:
1114:
1058:
1054:
1044:
1012:(4): 10–26.
1009:
1006:Acta Naturae
1005:
995:
955:(1): 35–51.
952:
948:
938:
890:(1): 14205.
887:
883:
873:
863:, retrieved
851:
828:, retrieved
816:
806:
797:
735:
731:
673:
669:
659:
616:
612:
602:
583:
577:
532:
528:
518:
455:
451:
441:
390:
386:
346:
338:
304:
295:
292:
288:
277:Deactivation
263:
246:
238:
235:Inactivation
226:
217:
208:
195:
184:
177:
170:
164:paddle model
163:
156:
149:
140:
131:
121:
100:
84:
57:
42:
38:ion channels
29:
23:
2076:(1): 1872.
676:(1): 1–24.
270:cytoplasmic
214:Cl Channels
192:Ca Channels
137:Na Channels
45:impermeable
28:, the term
2198:Categories
1822:2018-11-22
1734:(3): 207.
1350:: e36629.
1325:2023-04-27
865:2019-11-03
830:2019-10-29
817:StatPearls
330:References
146:K Channels
105:Activation
76:G proteins
34:activation
2166:2045-2322
2098:2041-1723
2033:0270-6474
1976:0022-3751
1919:0270-6474
1862:0022-1295
1750:1465-6906
1693:0896-6273
1637:0027-8424
1537:0007-1188
1480:0006-3002
1425:0031-9333
1366:2050-084X
1251:0007-1188
1192:1545-9985
1135:1943-0264
1075:1943-0264
969:0958-0670
912:2041-1723
752:0022-3751
690:0022-1295
633:0929-8673
492:0027-8424
415:0006-3495
68:phosphate
2184:30504856
2116:23695666
1994:21098008
1937:10024345
1817:30415410
1768:12620097
1709:41285799
1655:17101981
1498:20188062
1441:44165561
1433:29845874
1384:29809153
1320:35768555
1210:18552825
1153:22068972
1093:21746798
1036:25558391
987:24097157
930:28205548
860:22787615
825:30844170
770:21098008
708:26712848
651:23409712
569:10393876
510:11891285
433:28402882
313:See also
80:kinetics
2175:6269590
2146:Bibcode
2107:3674235
2078:Bibcode
2051:9133369
2042:6573675
1985:3055536
1928:6782169
1880:8882863
1871:2229322
1808:7086210
1701:8161454
1646:1693860
1615:Bibcode
1587:3315629
1546:3315629
1489:2885512
1375:6019066
1260:2884561
1201:2493294
1144:3249630
1084:3140680
1027:4273088
978:3885250
921:5316852
892:Bibcode
761:3055536
699:4692491
642:3706965
537:Bibcode
470:Bibcode
424:5389965
395:Bibcode
353:523–547
51:of the
2182:
2172:
2164:
2114:
2104:
2096:
2049:
2039:
2031:
1992:
1982:
1974:
1935:
1925:
1917:
1878:
1868:
1860:
1815:
1805:
1766:
1759:153452
1756:
1748:
1707:
1699:
1691:
1673:Neuron
1653:
1643:
1635:
1584:
1543:
1535:
1496:
1486:
1478:
1439:
1431:
1423:
1382:
1372:
1364:
1318:
1308:
1257:
1249:
1208:
1198:
1190:
1151:
1141:
1133:
1091:
1081:
1073:
1034:
1024:
985:
975:
967:
928:
918:
910:
858:
823:
768:
758:
750:
706:
696:
688:
649:
639:
631:
590:
567:
557:
508:
501:122561
498:
490:
431:
421:
413:
359:
30:gating
1705:S2CID
1437:S2CID
1344:eLife
560:33597
460:arXiv
205:cleft
2180:PMID
2162:ISSN
2112:PMID
2094:ISSN
2047:PMID
2029:ISSN
1990:PMID
1972:ISSN
1933:PMID
1915:ISSN
1876:PMID
1858:ISSN
1813:PMID
1764:PMID
1746:ISSN
1697:PMID
1689:ISSN
1651:PMID
1633:ISSN
1533:ISSN
1494:PMID
1476:ISSN
1464:1798
1429:PMID
1421:ISSN
1380:PMID
1362:ISSN
1316:PMID
1306:ISBN
1275:link
1247:ISSN
1206:PMID
1188:ISSN
1149:PMID
1131:ISSN
1089:PMID
1071:ISSN
1032:PMID
983:PMID
965:ISSN
926:PMID
908:ISSN
856:PMID
821:PMID
766:PMID
748:ISSN
704:PMID
686:ISSN
647:PMID
629:ISSN
588:ISBN
565:PMID
506:PMID
488:ISSN
429:PMID
411:ISSN
357:ISBN
264:The
183:The
169:The
162:The
155:The
53:cell
2170:PMC
2154:doi
2102:PMC
2086:doi
2037:PMC
2021:doi
1980:PMC
1964:doi
1960:589
1923:PMC
1907:doi
1866:PMC
1850:doi
1846:108
1803:PMC
1795:doi
1791:471
1754:PMC
1736:doi
1681:doi
1641:PMC
1623:doi
1611:103
1582:PMC
1574:doi
1570:164
1541:PMC
1525:doi
1521:164
1484:PMC
1468:doi
1411:doi
1370:PMC
1352:doi
1298:doi
1255:PMC
1239:doi
1235:158
1196:PMC
1180:doi
1139:PMC
1123:doi
1079:PMC
1063:doi
1022:PMC
1014:doi
973:PMC
957:doi
916:PMC
900:doi
756:PMC
740:doi
736:589
694:PMC
678:doi
674:147
637:PMC
621:doi
555:PMC
545:doi
496:PMC
478:doi
419:PMC
403:doi
391:112
24:In
2200::
2178:.
2168:.
2160:.
2152:.
2140:.
2136:.
2124:^
2110:.
2100:.
2092:.
2084:.
2072:.
2068:.
2045:.
2035:.
2027:.
2017:17
2015:.
2011:.
1988:.
1978:.
1970:.
1958:.
1954:.
1931:.
1921:.
1913:.
1903:19
1901:.
1897:.
1874:.
1864:.
1856:.
1844:.
1840:.
1811:.
1801:.
1789:.
1785:.
1762:.
1752:.
1744:.
1730:.
1726:.
1703:.
1695:.
1687:.
1677:12
1675:.
1663:^
1649:.
1639:.
1631:.
1621:.
1609:.
1605:.
1580:.
1568:.
1564:.
1539:.
1531:.
1519:.
1515:.
1492:.
1482:.
1474:.
1462:.
1458:.
1435:.
1427:.
1419:.
1407:98
1405:.
1401:.
1378:.
1368:.
1360:.
1346:.
1342:.
1314:,
1304:,
1292:,
1271:}}
1267:{{
1253:.
1233:.
1229:.
1218:^
1204:.
1194:.
1186:.
1176:15
1174:.
1170:.
1147:.
1137:.
1129:.
1117:.
1113:.
1101:^
1087:.
1077:.
1069:.
1057:.
1053:.
1030:.
1020:.
1008:.
1004:.
981:.
971:.
963:.
953:99
951:.
947:.
924:.
914:.
906:.
898:.
886:.
882:.
850:,
838:^
815:,
796:.
778:^
764:.
754:.
746:.
734:.
730:.
716:^
702:.
692:.
684:.
672:.
668:.
645:.
635:.
627:.
617:20
615:.
611:.
563:.
553:.
543:.
533:96
531:.
527:.
504:.
494:.
486:.
476:.
468:.
456:99
454:.
450:.
427:.
417:.
409:.
401:.
389:.
385:.
371:^
355:.
2186:.
2156::
2148::
2142:8
2118:.
2088::
2080::
2074:4
2053:.
2023::
1996:.
1966::
1939:.
1909::
1882:.
1852::
1825:.
1797::
1770:.
1738::
1732:4
1711:.
1683::
1657:.
1625::
1617::
1590:.
1576::
1549:.
1527::
1500:.
1470::
1443:.
1413::
1386:.
1354::
1348:7
1300::
1277:)
1263:.
1241::
1212:.
1182::
1155:.
1125::
1119:4
1095:.
1065::
1059:3
1038:.
1016::
1010:6
989:.
959::
932:.
902::
894::
888:8
800:.
772:.
742::
710:.
680::
653:.
623::
596:.
571:.
547::
539::
512:.
480::
472::
462::
435:.
405::
397::
365:.
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.