415:, females act more aggressively towards MHC-similar males than MHC-dissimilar males, suggesting the presence of female mate choice. Furthermore, males directed aggression at MHC-similar females. This was accompanied by male harassment of unreceptive females; however, there was a positive correlation between male aggression and reproductive success. The ability of the males to over-power the females' original mate choice resulted in the offspring of the targets of male aggression having low genetic diversity. Offspring with high genetic diversity seemed to happen only when the operational sex ratio was female-biased, when females were more likely to be able to exert mate choice, and males were less likely to harass females. These results suggest that sexual conflict may interfere with female mate choice for 'good' MHC genes.
275:
would be expected by chance. If the selection for shirts was not random, and actually selected for MHC-dissimilar alleles, this suggests that MHC genetic composition does influence mate choice. Furthermore, when the degree of similarity between the wearer and the smeller was statistically accounted for, there was no longer a significant influence of MHC on odor preference. The results show that MHC similarity or dissimilarity certainly plays a role in mate choice. Specifically, MHC-disassortative mate choice and less similar MHC combinations are selected for. One interesting aspect of the
Wedekind's experiment was that in contrast to normally cycling women, women taking oral contraceptives preferred odors of MHC-similar men. This would suggest that the pill may interfere with the adaptive preference for dissimilarity.
307:, by observing effects of MHC upon natural spawning salmon that resided in the river versus artificial crosses that were carried out in hatcheries. Logically, the artificial crosses would be bereft of the benefits of mate choice that would naturally be available. The results showed that the offspring of the artificially bred salmon were more infected with parasites: almost four times more than the naturally-spawned offspring were. In addition, wild offspring were more MHC-heterozygous than the artificially-bred offspring. These results support the Heterozygous Advantage hypothesis of sexual selection for MHC-dissimilar mate choice. In another fish, the three-spined stickleback, it has been shown that females desire MHC diversity in their offspring, which affects their mate choice.
147:
would be to avoid mating with family members and any harmful genetic consequences that could occur as a result. The hypothesis states that inbreeding increases the amount of overall homozygosity—not just locally in the MHC, so an increase in genetic homozygosity may be accompanied not only by the expression of recessive diseases and mutations, but by the loss of any potential heterozygote advantage as well. Animals only rarely avoid inbreeding. The inbreeding avoidance hypothesis has been "ruled out as an explanation for the observed pattern of MHC-dependent mate preference" because relatedness is not associated with mate choice.
367:. In this case, however, it seems that sperm selection is more dependent on the ovum. MHC-heterozygous males were found to have significantly more fertilization success than MHC-homozygous males; sperm count, motility, and swimming velocity were not shown to significantly co-vary with similarity or dissimilarity at the MHC. It is proposed that there is a chemo-attraction system responsible for the egg itself being able to discriminate and selectively choose between MHC-heterozygous and MHC-homozygous males.
235:
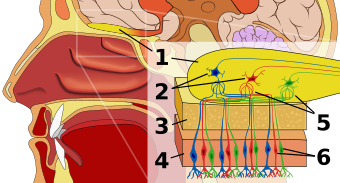
individual. During cellular turnover, the MHC-peptide complex is shed from the cell surface and the fragments are dispensed in bodily fluids such as blood serum, saliva, and urine. Scientists believe that commensal microflora, microorganisms that line epithelial surfaces open to the external environment such as the gastrointestinal tract and vagina, further degrade these fragments, which are made volatile by this process. Recently, it has been shown that receptors in the
20:
107:
associated with MHC loci. In humans, those with greater MHC diversity have a greater risk for autoimmune disorders. MHC diversity may be low "because foreign peptides have to stand out against the self-background." On an individual level, MHC diversity tends to be low. Across many species, there is intermediate heterozygosity in the MHC. Overall evidence supports intermediate MHC heterozygosity is best.
196:
123:
increases. An advantageous characteristic that allows a parasite to escape recognition spreads, and causes selection against what was formerly a resistant allele. This enables the parasite to escape this cycle of frequency-dependent selection, and such a cycle eventually leads to a co-evolutionary arms race that may support the maintenance of MHC diversity. This hypothesis has empirical support.
183:
females to be able to choose a male that provided either an indirect or direct benefit. As a result of female choice, sexual selection is imposed on males. This is evidenced by genetic "advertisement"—an example of this would be the existence of exaggerated traits, such as the elaborate tail-feathers of male peacocks. However, in humans, both sexes exert mate choice.
127:
175:
289:, post-copulatory mate-choice is associated with genetic constitution. Fathers are more MHC-dissimilar from the mother than are randomly tested males. Fathers have more differences in amino acid and microsatellite diversity than did randomly tested males. It is hypothesized that this is caused by female cryptic choice.
358:, is one species in which sperm competition is influenced by the variation in the major histocompatibility complex, specifically that of the Class I alleles. Atlantic salmon males have higher rates of successful fertilization when competing for eggs from females genetically similar at the class I genes of the MHC.
234:
Although it is not known exactly how MHC-specific odors are recognized, it is currently believed that proteins bound to the peptide-binding groove of the MHC may produce the odorant. Each MHC protein binds to a specific peptide sequence, yielding a set of uniquely bound peptide-MHC complexes for each
106:
The optimality hypothesis states too much variability in the MHC can result in a failure of T-cells to distinguish themselves non-selves, and thereby increase the risk of autoimmune disease. This would confer greater fitness to individuals without a large degree MHC diversity. Autoimmune diseases are
53:
affecting an organism's fitness stems from the co-evolutionary arms race between hosts and parasites. There are two hypotheses for explaining the MHC's high diversity, which are not mutually exclusive. One is that there is selection for individuals to possess a diverse set of MHC alleles, which would
170:
in some species. Depending on how parasites alter selection on MHC alleles, MHC-dependent mate-choice may increase the fitness of the offspring by enhancing its immunity, as mentioned earlier. If this is the case, either through the heterozygote advantage hypothesis or the Red Queen hypothesis, then
146:
hypothesis has less to do with host-parasite relationships than does the heterozygote advantage hypothesis or the Red Queen hypothesis. The extreme diversity in the MHC would cause individuals sharing MHC alleles to be more likely to be related. As a result, one function of MHC-disassortative mating
386:
behave similarly to the male junglefowl. Initial copulation between a male and a female without any rivals was shown to be extended when the male sensed a higher female fecundity. However, second males adjusted the duration of their copulation depending on the relatedness between the female and the
274:
In another study done by
Wedekind et al., 121 women and men were asked to rank the pleasantness of the odors of sweaty T-shirts. Upon smelling the shirts, it was found that men and women who were reminded of their own mate or ex-mate had dramatically fewer MHC alleles in common with the wearer than
191:
MHC-based sexual selection is known to involve olfactory mechanisms in such vertebrate taxa as fish, mice, humans, primates, birds, and reptiles. At its simplest level, humans have long been acquainted with the sense of olfaction for its use in determining the pleasantness or the unpleasantness of
102:
advantage. Unfortunately, the MHC-heterozygote advantage hypothesis has not been adequately tested. A non-MHC immune genes across species exhibit heterozygote disadvantage, or no advantage. In mice, increased MHC heterozygosity reduces fitness, challenging this hypothesis. MHC-heterozygous females
270:
Other studies have approached mate choice based on odor preference. In one study done by
Wedekind et al., women were asked to smell male axillary odors collected on T-shirts worn by different males. Women that were ovulating rated the odors of MHC-dissimilar men as more pleasant than those of the
226:
Chemosensation, which is one of the most primitive senses, has evolved into a specialized sensory system. Humans can not only detect, but also assess, and respond to environmental (chemical) olfactory cues—especially those used to evoke behavioral and sexual responses from other individuals, also
182:
Therefore, mate choice—with respect to the MHC—has probably evolved so that females choose males either based on diverse genes (heterozygote advantage and inbreeding avoidance hypotheses) or "good" genes. The fact that females choose is naturally selected, as it would be an advantageous trait for
390:
Abortional selection may be a form of cryptic female choice. Many studies on humans and rodents have found that females may spontaneously abort pregnancies in which the offspring is too MHC-similar. In addition, in vitro fertilizations are more likely to fail when couples have similar MHC genes.
334:
Even though many species are socially monogamous, females can accept or actively seek mating outside of the relationship; extra-pair paternity is a mating pattern known to be affiliated with MHC-associated mate choice. Birds are one of the more commonly studied groups of animals to exhibit this
69:
There is evidence that many vertebrates, including humans, select their mates based on signals of "compatibility" between their MHC alleles, with a preference for mates with different alleles than their own, resulting in pairings that would tend to produce more heterozygous offspring. There are
122:
cause generation of new variants among offspring, which may facilitate a quick response to rapidly evolving parasites or pathogens with much shorter generation times. However, if this particular allele becomes common, selection pressure on parasites to avoid recognition by this common allele
376:
males instead of females exert cryptic preference. Male junglefowl showed no preference when simultaneously presented with both an MHC-dissimilar and an MHC-similar female. However, they did show a cryptic preference by allocating more sperm to the more MHC-dissimilar of the two.
266:
community and found dramatically fewer HLA matches between husbands and wives than expected when considering the social structure of their community. On the other hand, there was no evidence of MHC-based mate choice in the same study of 200 couples from South
Amerindian tribes.
252:
MHC similarity in humans has been studied in three broad ways: odor, facial attractiveness, and actual mate choice. Studies of odor find MHC-dissimilarity preferences but vary in details, while facial attractiveness favors MHC-similarity and actual mating studies are varied.
347:, there was no evidence of MHC variation between social mates. However, when females' social mates were MHC-similar, they were more likely to participate in extra-pair paternity; in most cases, the extra-pair male was significantly more MHC-dissimilar than the social mate.
297:
In mice, both males and females choose MHC-dissimilar partners. Mice develop the ability to identify family members during early growth and are known to avoid inbreeding with kin, which would support the MHC-mediated mate choice hypothesis for inbreeding avoidance.
261:
Several studies suggest that MHC-related odor preferences and mate choice are demonstrated by humans. However, the role of MHC in human mate choice has been relatively controversial. One study conducted by Ober et al. examined HLA types from 400 couples in the
154:
occurs for "good" genes, then it is implied that genetic variation exists among males. Furthermore, one would presume that said difference in genes would impart a difference in fitness as well, which could potentially be chosen or selected for.
387:
first male, believed to be determined by the MHC-odor of the copulatory plug. A closer genetic relatedness between a male and a female sand lizard increased the chances for a successful fertilization and rate of paternity for the second male.
318:, chose MHC-dissimilar males to mate with. Females are more likely to engage in extra-pair relationships if paired with MHC-similar mates and more dissimilar mates are available. Similarly, MHC diversity in house sparrows,
34:(MHC) genes code for cell surface proteins that facilitate an organism's immune response to pathogens as well as its ability to avoid attacking its own cells. These genes have maintained an unusually high level of
231:. Pheromones function to communicate one's species, sex, and perhaps most importantly one's genetic identity. The genes of the MHC provide the basis from which a set of unique olfactory coding develops.
103:
had significantly reduced fitness compared to homozygotes. This finding has been replicated in another study in mice and again in fish In some cases, excess heterozygosity can lead to decreased fitness.
271:
MHC-similar men. Furthermore, odors of MHC-dissimilar men often reminded women of current or former partners, suggesting that odor—specifically odor for MHC-dissimilarity—plays a role in mate choice.
239:
of mice are activated by peptides having similar characteristics to MHC proteins; further studies may hopefully soon clarify the exact transformation between MHC genotype and an olfactory mechanism.
38:
diversity throughout time and throughout different populations. This means that for each MHC gene, many alleles (or gene variants) consistently exist within the population, and many individuals are
150:
In the course of searching for potential mates, it would benefit females to be able to discriminate against "bad" genes in order to increase the health and viability of their offspring. If female
70:
several proposed hypotheses that address how MHC-associated mating preferences could be adaptive and how an unusually large amount of allelic diversity has been maintained in the MHC.
114:
render different resistances to a particular parasite, then the allele with the highest resistance is favored, selected for, and consequently spread throughout the population.
429:
405:
If males attempt to thwart female mate choice by mating with a female against her will, sexual conflict may interfere with the choice for compatibility at the MHC genes.
705:"Naturally occurring Toll-like receptor 11 (TLR11) and Toll-like receptor 12 (TLR12) polymorphisms are not associated with Toxoplasma gondii infection in wild wood mice"
1038:
192:
one's resources, food, etc. At a deeper level, it has been predicted that olfaction serves to personally identify individuals based upon the genes of the MHC.
90:, then it is beneficial for females to choose mates with MHC genes different from their own, and would result in MHC-heterozygous offspring—this is known as
703:
Morger, Jennifer; Bajnok, Jaroslav; Boyce, Kellyanne; Craig, Philip S.; Rogan, Michael T.; Lun, Zhao-Rong; Hide, Geoff; Tschirren, Barbara (2014-08-01).
537:
O'Dwyer TW, Nevitt GA (July 2009). "Individual odor recognition in procellariiform chicks: potential role for the major histocompatibility complex".
581:
Quéméré, Erwan; Rossi, Sophie; Petit, Elodie; Marchand, Pascal; Merlet, Joël; Game, Yvette; Galan, Maxime; Gilot-Fromont, Emmanuelle (2020-03-10).
1269:"Major histocompatibility complex variability in the clonal Amazon molly, Poecilia formosa : is copy number less important than genotype?"
1130:
1014:
1389:
Westemeier RL, Brawn JD, Simpson SA, et al. (November 1998). "Tracking the long-term decline and recovery of an isolated population".
2324:
Olsson M, Madsen T, Ujvari B, Wapstra E (April 2004). "Fecundity and MHC affects ejaculation tactics and paternity bias in sand lizards".
350:
MHC-mediated mate choice may occur after copulation, at the gametic level, through sperm competition or female cryptic choice. The
341:, females engaged in extra-pair paternity much less frequently when their mates were MHC-heterozygous. In the Seychelles warbler
31:
2125:
Promerová Vinkler. Occurrence of extra-pair paternity is connected to social male's MHC-variability in the scarlet rosefinch
897:
Nelson-Flower, Martha J; Germain, Ryan R; MacDougall-Shackleton, Elizabeth A; Taylor, Sabrina S; Arcese, Peter (2018-06-27).
1564:
472:
301:
Fish are another group of vertebrates shown to display MHC-associated mate choice. Scientists tested the
Atlantic salmon,
24:
946:
Ilmonen, Petteri; Penn, Dustin J; Damjanovich, Kristy; Morrison, Linda; Ghotbi, Laleh; Potts, Wayne K (2007-08-01).
400:
2381:"Sexual conflict inhibits female mate choice for major histocompatibility complex dissimilarity in Chinook salmon"
1324:"MHC genes and parasitism in Carassius gibelio, a diploid-triploid fish species with dual reproduction strategies"
94:. The hypothesis states that individuals with a heterozygous MHC would be capable of recognizing a wider range of
2440:
343:
331:. Females preferred to associate with odor samples obtained from males more distantly related at the MHC I loci.
1978:"Major histocompatibility complex diversity influences parasite resistance and innate immunity in sticklebacks"
1828:"Body odour preferences in men and women: do they aim for specific MHC combinations or simply heterozygosity?"
2435:
2239:
Skarstein F, et al. (2005). "MHC and fertilization success in the Arctic charr (Salvelinus alpinus)".
753:
Antonides, Jennifer; Mathur, Samarth; Sundaram, Mekala; Ricklefs, Robert; DeWoody, J. Andrew (2019-05-22).
440:
216:
98:
and therefore of inciting a specific immune response against a greater number of pathogens—thus having an
56:
337:
1469:
539:
115:
91:
1268:
704:
648:"Selection Balancing at Innate Immune Genes: Adaptive Polymorphism Maintenance in Toll-Like Receptors"
60:. The second is that there is selection that undergoes a frequency-dependent cycle—this is called the
2191:"Atlantic salmon eggs favour sperm in competition that have similar major histocompatibility alleles"
1572:
1400:
1335:
1280:
1164:
837:
766:
594:
480:
163:
159:
143:
62:
110:
The Red Queen hypothesis asserts that MHC diversity is maintained by parasites. If individuals' MHC
2326:
2078:"Reed bunting females increase fitness through extra-pair mating with genetically dissimilar males"
583:"Genetic epidemiology of the Alpine ibex reservoir of persistent and virulent brucellosis outbreak"
2361:
2256:
1697:
1619:
1535:
1457:
1424:
1304:
1032:
948:"Major Histocompatibility Complex Heterozygosity Reduces Fitness in Experimentally Infected Mice"
879:
563:
363:
285:
220:
99:
1498:"MHC studies in nonmodel vertebrates: what have we learned about natural selection in 15 years?"
1444:"Faculty Opinions recommendation of Meta-analytic evidence that animals rarely avoid inbreeding"
2411:
2353:
2306:
2221:
2171:
2108:
2058:
2008:
1958:
1908:
1858:
1805:
1752:
1689:
1651:
1600:
1527:
1519:
1416:
1391:
1371:
1353:
1296:
1249:
1200:
1182:
1126:
1091:
1073:
1054:"Major Histocompatibility Complex-Dependent Susceptibility to Cryptococcus neoformans in Mice"
1020:
1010:
985:
967:
928:
920:
871:
863:
802:
784:
732:
724:
685:
667:
628:
610:
555:
508:
236:
167:
50:
19:
2401:
2393:
2385:
2343:
2335:
2296:
2288:
2280:
2248:
2211:
2203:
2195:
2161:
2153:
2145:
2098:
2090:
2082:
2048:
2040:
2032:
1998:
1990:
1982:
1948:
1940:
1932:
1898:
1890:
1882:
1848:
1840:
1832:
1795:
1787:
1779:
1742:
1732:
1681:
1641:
1633:
1590:
1580:
1509:
1447:
1408:
1361:
1343:
1288:
1239:
1231:
1190:
1172:
1118:
1081:
1065:
975:
959:
910:
853:
845:
792:
774:
716:
675:
659:
618:
602:
547:
498:
488:
311:
43:
283:
There is evidence of MHC-associated mate choice in other primates. In the grey mouse lemur
54:
occur if MHC heterozygotes are more resistant to pathogens than homozygotes—this is called
1482:
468:"Mate choice decisions of stickleback females predictably modified by MHC peptide ligands"
351:
1576:
1404:
1339:
1284:
1168:
841:
821:
770:
598:
484:
2406:
2380:
2301:
2275:
2216:
2190:
2166:
2140:
2103:
2077:
2053:
2027:
2003:
1977:
1953:
1927:
1903:
1877:
1853:
1827:
1800:
1774:
1747:
1719:
1646:
1366:
1323:
1244:
1219:
980:
797:
754:
680:
623:
582:
466:
Milinski M, Griffiths S, Wegner KM, Reusch TB, Haas-Assenbaum A, Boehm T (March 2005).
409:
382:
327:
200:
2274:
Gillingham MA, Richardson DS, Løvlie H, Moynihan A, Worley K, Pizzari T (March 2009).
1220:"Does intra-individual major histocompatibility complex diversity keep a golden mean?"
1086:
1053:
820:
Cornetti, Luca; Hilfiker, Daniela; Lemoine, Mélissa; Tschirren, Barbara (2018-08-06).
503:
467:
2429:
2365:
2348:
2276:"Cryptic preference for MHC-dissimilar females in male red junglefowl, Gallus gallus"
1672:
Havlicek J, Roberts SC (May 2009). "MHC-correlated mate choice in humans: A review".
1595:
1559:
1514:
1497:
1461:
1292:
1195:
1152:
551:
372:
2260:
1701:
1539:
1428:
1322:Šimková, Andrea; Košař, Martin; Vetešník, Lukáš; Vyskočilová, Martina (2013-06-14).
1308:
1069:
883:
567:
2141:"MHC-based patterns of social and extra-pair mate choice in the Seychelles warbler"
1685:
1153:"The optimal number of major histocompatibility complex molecules in an individual"
899:"Purifying Selection in the Toll-Like Receptors of Song Sparrows Melospiza melodia"
204:
79:
1452:
1443:
1412:
1737:
1122:
720:
178:
The exaggerated, elongated upper tail coverts make up the "train" of the peacock.
1878:"Compatibility counts: MHC-associated mate choice in a wild promiscuous primate"
1724:
1624:
1218:
Woelfing, Benno; Traulsen, Arne; Milinski, Manfred; Boehm, Thomas (2009-01-12).
963:
361:
Another species that exhibits MHC-associated cryptic choice is the Arctic charr
303:
151:
2026:
Olsson M, Madsen T, Nordby J, Wapstra E, Ujvari B, Wittsell H (November 2003).
1775:"MHC-correlated odour preferences in humans and the use of oral contraceptives"
1637:
1110:
606:
195:
2252:
947:
779:
212:
131:
87:
1523:
1357:
1348:
1186:
1177:
1077:
1024:
971:
924:
867:
788:
728:
671:
663:
614:
1111:"Polymorphism at MHC Loci and Isolation by the Immune System in Vertebrates"
915:
898:
755:"Immunogenetic response of the bananaquit in the face of malarial parasites"
493:
434:
424:
263:
228:
2415:
2397:
2357:
2310:
2292:
2225:
2207:
2175:
2157:
2112:
2094:
2062:
2044:
2012:
1994:
1962:
1944:
1912:
1894:
1844:
1809:
1791:
1756:
1693:
1655:
1585:
1531:
1375:
1300:
1253:
1235:
1095:
989:
932:
875:
806:
736:
689:
632:
559:
512:
1862:
1604:
1420:
1204:
822:"Small-scale spatial variation in infection risk shapes the evolution of a
325:
MHC-mediated mate choice has been shown to exist in
Swedish sand lizards,
126:
647:
119:
95:
83:
39:
1052:
McClelland, Erin E.; Granger, Donald L.; Potts, Wayne K. (August 2003).
858:
1558:
Yamazaki K, Beauchamp GK, Singer A, Bard J, Boyse EA (February 1999).
1224:
Philosophical
Transactions of the Royal Society B: Biological Sciences
849:
46:(meaning they possess two different alleles for a given gene locus).
1004:
111:
35:
370:
Contrary to the
Atlantic salmon and the Arctic char, red junglefowl
174:
2339:
2028:"Major histocompatibility complex and mate choice in sand lizards"
1928:"MHC-mediated mate choice increases parasite resistance in salmon"
194:
173:
125:
18:
208:
135:
2139:
Richardson DS, Komdeur J, Burke T, von
Schantz T (April 2005).
171:
selection also favors mating practices that are MHC-dependent.
1976:
Kurtz J, Kalbe M, Aeschlimann PB, et al. (January 2004).
162:
of MHC genes is selected for by host-parasite arms races (the
2189:
Yeates SE, Einum S, Fleming IA, et al. (February 2009).
1773:
Roberts SC, Gosling LM, Carter V, Petrie M (December 2008).
2379:
Garner SR, Bortoluzzi RN, Heath DD, Neff BD (March 2010).
2076:
Suter SM, Keiser M, Feignoux R, Meyer DR (November 2007).
1151:
Nowak, M A; Tarczy-Hornoch, K; Austyn, J M (1992-11-15).
1267:
Lampert, K. P.; Fischer, P.; Schartl, M. (March 2009).
322:, suggests that MHC-disassortative mate choice occurs.
130:
Parasites are in a constant arms race with their host:
166:); however, disassortative mate choice may maintain
1442:Schlupp, Ingo; Berbel-Filho, Waldir (2021-06-03).
1115:Non-Neutral Evolution: Theories and Molecular Data
430:Body odor and subconscious human sexual attraction
1109:Takahata, Naoyuki (1994), Golding, Brian (ed.),
1157:Proceedings of the National Academy of Sciences
748:
746:
1876:Schwensow N, Eberle M, Sommer S (March 2008).
1620:"Sex and the nose: human pheromonal responses"
1003:Joe., Demas, Gregory E. Nelson, Randy (2012).
23:MHC sexual selection has been observed in the
1926:Consuegra S, Garcia de Leaniz C (June 2008).
1117:, Boston, MA: Springer US, pp. 233–246,
646:Minias, Piotr; Vinkler, Michal (2022-05-01).
8:
1037:: CS1 maint: multiple names: authors list (
1821:
1819:
1553:
1551:
1549:
1768:
1766:
461:
459:
457:
335:sexual behavior. In the scarlet rosefinch
187:The relationship between olfaction and MHC
2405:
2347:
2300:
2215:
2165:
2102:
2052:
2002:
1952:
1902:
1852:
1799:
1746:
1736:
1720:"Is mate choice in humans MHC-dependent?"
1645:
1594:
1584:
1560:"Odortypes: their origin and composition"
1513:
1451:
1365:
1347:
1243:
1194:
1176:
1085:
979:
914:
857:
796:
778:
679:
622:
502:
492:
2129:Journal of Avian Biology 42, 5-10(2011).
1713:
1711:
1009:. Oxford University Press. p. 238.
78:In the first hypothesis, if individuals
1496:Bernatchez, L.; Landry, C. (May 2003).
532:
530:
528:
526:
524:
522:
453:
1478:
1467:
1030:
1667:
1665:
7:
2241:Behavioral Ecology and Sociobiology
1826:Wedekind C, Füri S (October 1997).
1718:Chaix R, Cao C, Donnelly P (2008).
14:
709:Infection, Genetics and Evolution
82:at the MHC are more resistant to
1515:10.1046/j.1420-9101.2003.00531.x
1293:10.1111/j.1365-294X.2009.04097.x
826:resistance gene in wild rodents"
552:10.1111/j.1749-6632.2009.03887.x
32:Major histocompatibility complex
1502:Journal of Evolutionary Biology
1070:10.1128/IAI.71.8.4815-4817.2003
652:Molecular Biology and Evolution
1686:10.1016/j.psyneuen.2008.10.007
16:Adaptive immune gene selection
1:
1565:Proc. Natl. Acad. Sci. U.S.A.
1453:10.3410/f.740048135.793586159
1413:10.1126/science.282.5394.1695
473:Proc. Natl. Acad. Sci. U.S.A.
1738:10.1371/journal.pgen.1000184
1123:10.1007/978-1-4615-2383-3_19
721:10.1016/j.meegid.2014.05.032
964:10.1534/genetics.107.074815
199:Human olfactory system. 1:
25:black-throated blue warbler
2457:
1638:10.1177/014107680710000612
607:10.1038/s41598-020-61299-2
401:Interlocus sexual conflict
398:
2253:10.1007/s00265-004-0860-z
780:10.1186/s12862-019-1435-y
344:Acrocephalus sechellensis
316:Passerculus sandwichensis
1674:Psychoneuroendocrinology
1349:10.1186/1471-2148-13-122
1328:BMC Evolutionary Biology
1178:10.1073/pnas.89.22.10896
759:BMC Evolutionary Biology
221:Olfactory receptor cells
1618:Bhutta MF (June 2007).
494:10.1073/pnas.0408264102
413:Oncorhyncus tshawytscha
395:MHC and sexual conflict
158:Generally, the extreme
2398:10.1098/rspb.2009.1639
2293:10.1098/rspb.2008.1549
2208:10.1098/rspb.2008.1257
2158:10.1098/rspb.2004.3028
2127:Carpodacus erythrinus.
2095:10.1098/rspb.2007.0799
2045:10.1098/rsbl.2003.0079
1995:10.1098/rspb.2003.2567
1945:10.1098/rspb.2008.0066
1895:10.1098/rspb.2007.1433
1845:10.1098/rspb.1997.0204
1792:10.1098/rspb.2008.0825
1586:10.1073/pnas.96.4.1522
1477:Cite journal requires
1236:10.1098/rstb.2008.0174
1058:Infection and Immunity
664:10.1093/molbev/msac102
441:The Compatibility Gene
223:
217:Glomerulus (olfaction)
179:
139:
57:heterozygote advantage
28:
2349:10536/DRO/DU:30066491
916:10.1093/jhered/esy027
540:Ann. N. Y. Acad. Sci.
198:
177:
129:
92:disassortative mating
22:
164:Red Queen hypothesis
144:inbreeding avoidance
86:than those that are
63:Red Queen hypothesis
2039:(Suppl 2): S254–6.
1577:1999PNAS...96.1522Y
1405:1998Sci...282.1695W
1340:2013BMCEE..13..122S
1285:2009MolEc..18.1124L
1169:1992PNAS...8910896N
1163:(22): 10896–10899.
903:Journal of Heredity
842:2018MolEc..27.3515C
771:2019BMCEE..19..107A
599:2020NatSR..10.4400Q
485:2005PNAS..102.4414M
338:Carpocus erythrinus
49:The vast source of
1939:(1641): 1397–403.
587:Scientific Reports
380:Male sand lizards
364:Salvelinus alpinus
286:Microcebus murinus
243:Empirical evidence
224:
180:
140:
29:
2287:(1659): 1083–92.
2089:(1627): 2865–71.
1989:(1535): 197–204.
1786:(1652): 2715–22.
1273:Molecular Ecology
1230:(1513): 117–128.
1132:978-1-4615-2383-3
1016:978-0-19-987624-2
850:10.1111/mec.14812
836:(17): 3515–3524.
830:Molecular Ecology
320:Passer domesticus
312:Savannah sparrows
237:vomeronasal organ
168:genetic diversity
51:genetic variation
2448:
2441:Sexual selection
2420:
2419:
2409:
2392:(1683): 885–94.
2386:Proc. Biol. Sci.
2376:
2370:
2369:
2351:
2321:
2315:
2314:
2304:
2281:Proc. Biol. Sci.
2271:
2265:
2264:
2236:
2230:
2229:
2219:
2202:(1656): 559–66.
2196:Proc. Biol. Sci.
2186:
2180:
2179:
2169:
2152:(1564): 759–67.
2146:Proc. Biol. Sci.
2136:
2130:
2123:
2117:
2116:
2106:
2083:Proc. Biol. Sci.
2073:
2067:
2066:
2056:
2033:Proc. Biol. Sci.
2023:
2017:
2016:
2006:
1983:Proc. Biol. Sci.
1973:
1967:
1966:
1956:
1933:Proc. Biol. Sci.
1923:
1917:
1916:
1906:
1889:(1634): 555–64.
1883:Proc. Biol. Sci.
1873:
1867:
1866:
1856:
1839:(1387): 1471–9.
1833:Proc. Biol. Sci.
1823:
1814:
1813:
1803:
1780:Proc. Biol. Sci.
1770:
1761:
1760:
1750:
1740:
1715:
1706:
1705:
1669:
1660:
1659:
1649:
1615:
1609:
1608:
1598:
1588:
1555:
1544:
1543:
1517:
1493:
1487:
1486:
1480:
1475:
1473:
1465:
1455:
1439:
1433:
1432:
1399:(5394): 1695–8.
1386:
1380:
1379:
1369:
1351:
1319:
1313:
1312:
1279:(6): 1124–1136.
1264:
1258:
1257:
1247:
1215:
1209:
1208:
1198:
1180:
1148:
1142:
1141:
1140:
1139:
1106:
1100:
1099:
1089:
1064:(8): 4815–4817.
1049:
1043:
1042:
1036:
1028:
1000:
994:
993:
983:
958:(4): 2501–2508.
943:
937:
936:
918:
894:
888:
887:
861:
817:
811:
810:
800:
782:
750:
741:
740:
700:
694:
693:
683:
643:
637:
636:
626:
578:
572:
571:
534:
517:
516:
506:
496:
463:
293:In other animals
257:Specific studies
2456:
2455:
2451:
2450:
2449:
2447:
2446:
2445:
2426:
2425:
2424:
2423:
2378:
2377:
2373:
2323:
2322:
2318:
2273:
2272:
2268:
2238:
2237:
2233:
2188:
2187:
2183:
2138:
2137:
2133:
2124:
2120:
2075:
2074:
2070:
2025:
2024:
2020:
1975:
1974:
1970:
1925:
1924:
1920:
1875:
1874:
1870:
1825:
1824:
1817:
1772:
1771:
1764:
1731:(9): e1000184.
1717:
1716:
1709:
1671:
1670:
1663:
1617:
1616:
1612:
1557:
1556:
1547:
1495:
1494:
1490:
1476:
1466:
1441:
1440:
1436:
1388:
1387:
1383:
1321:
1320:
1316:
1266:
1265:
1261:
1217:
1216:
1212:
1150:
1149:
1145:
1137:
1135:
1133:
1108:
1107:
1103:
1051:
1050:
1046:
1029:
1017:
1002:
1001:
997:
945:
944:
940:
896:
895:
891:
819:
818:
814:
752:
751:
744:
702:
701:
697:
645:
644:
640:
580:
579:
575:
536:
535:
520:
465:
464:
455:
450:
421:
403:
397:
352:Atlantic salmon
295:
281:
259:
250:
245:
189:
134:suffering from
76:
17:
12:
11:
5:
2454:
2452:
2444:
2443:
2438:
2436:Histochemistry
2428:
2427:
2422:
2421:
2371:
2340:10.1554/03-610
2316:
2266:
2247:(4): 374–380.
2231:
2181:
2131:
2118:
2068:
2018:
1968:
1918:
1868:
1815:
1762:
1707:
1680:(4): 497–512.
1661:
1610:
1545:
1508:(3): 363–377.
1488:
1479:|journal=
1434:
1381:
1314:
1259:
1210:
1143:
1131:
1101:
1044:
1015:
995:
938:
909:(5): 501–509.
889:
812:
742:
695:
658:(5): msac102.
638:
573:
518:
479:(12): 4414–8.
452:
451:
449:
446:
445:
444:
437:
432:
427:
420:
417:
410:Chinook salmon
396:
393:
383:Lacerta agilis
328:Lacerta agilis
294:
291:
280:
277:
258:
255:
249:
246:
244:
241:
201:Olfactory bulb
188:
185:
75:
72:
15:
13:
10:
9:
6:
4:
3:
2:
2453:
2442:
2439:
2437:
2434:
2433:
2431:
2417:
2413:
2408:
2403:
2399:
2395:
2391:
2388:
2387:
2382:
2375:
2372:
2367:
2363:
2359:
2355:
2350:
2345:
2341:
2337:
2333:
2329:
2328:
2320:
2317:
2312:
2308:
2303:
2298:
2294:
2290:
2286:
2283:
2282:
2277:
2270:
2267:
2262:
2258:
2254:
2250:
2246:
2242:
2235:
2232:
2227:
2223:
2218:
2213:
2209:
2205:
2201:
2198:
2197:
2192:
2185:
2182:
2177:
2173:
2168:
2163:
2159:
2155:
2151:
2148:
2147:
2142:
2135:
2132:
2128:
2122:
2119:
2114:
2110:
2105:
2100:
2096:
2092:
2088:
2085:
2084:
2079:
2072:
2069:
2064:
2060:
2055:
2050:
2046:
2042:
2038:
2035:
2034:
2029:
2022:
2019:
2014:
2010:
2005:
2000:
1996:
1992:
1988:
1985:
1984:
1979:
1972:
1969:
1964:
1960:
1955:
1950:
1946:
1942:
1938:
1935:
1934:
1929:
1922:
1919:
1914:
1910:
1905:
1900:
1896:
1892:
1888:
1885:
1884:
1879:
1872:
1869:
1864:
1860:
1855:
1850:
1846:
1842:
1838:
1835:
1834:
1829:
1822:
1820:
1816:
1811:
1807:
1802:
1797:
1793:
1789:
1785:
1782:
1781:
1776:
1769:
1767:
1763:
1758:
1754:
1749:
1744:
1739:
1734:
1730:
1727:
1726:
1721:
1714:
1712:
1708:
1703:
1699:
1695:
1691:
1687:
1683:
1679:
1675:
1668:
1666:
1662:
1657:
1653:
1648:
1643:
1639:
1635:
1632:(6): 268–74.
1631:
1627:
1626:
1621:
1614:
1611:
1606:
1602:
1597:
1592:
1587:
1582:
1578:
1574:
1571:(4): 1522–5.
1570:
1567:
1566:
1561:
1554:
1552:
1550:
1546:
1541:
1537:
1533:
1529:
1525:
1521:
1516:
1511:
1507:
1503:
1499:
1492:
1489:
1484:
1471:
1463:
1459:
1454:
1449:
1445:
1438:
1435:
1430:
1426:
1422:
1418:
1414:
1410:
1406:
1402:
1398:
1394:
1393:
1385:
1382:
1377:
1373:
1368:
1363:
1359:
1355:
1350:
1345:
1341:
1337:
1333:
1329:
1325:
1318:
1315:
1310:
1306:
1302:
1298:
1294:
1290:
1286:
1282:
1278:
1274:
1270:
1263:
1260:
1255:
1251:
1246:
1241:
1237:
1233:
1229:
1225:
1221:
1214:
1211:
1206:
1202:
1197:
1192:
1188:
1184:
1179:
1174:
1170:
1166:
1162:
1158:
1154:
1147:
1144:
1134:
1128:
1124:
1120:
1116:
1112:
1105:
1102:
1097:
1093:
1088:
1083:
1079:
1075:
1071:
1067:
1063:
1059:
1055:
1048:
1045:
1040:
1034:
1026:
1022:
1018:
1012:
1008:
1007:
1006:Ecoimmunology
999:
996:
991:
987:
982:
977:
973:
969:
965:
961:
957:
953:
949:
942:
939:
934:
930:
926:
922:
917:
912:
908:
904:
900:
893:
890:
885:
881:
877:
873:
869:
865:
860:
855:
851:
847:
843:
839:
835:
831:
827:
825:
816:
813:
808:
804:
799:
794:
790:
786:
781:
776:
772:
768:
764:
760:
756:
749:
747:
743:
738:
734:
730:
726:
722:
718:
714:
710:
706:
699:
696:
691:
687:
682:
677:
673:
669:
665:
661:
657:
653:
649:
642:
639:
634:
630:
625:
620:
616:
612:
608:
604:
600:
596:
592:
588:
584:
577:
574:
569:
565:
561:
557:
553:
549:
545:
542:
541:
533:
531:
529:
527:
525:
523:
519:
514:
510:
505:
500:
495:
490:
486:
482:
478:
475:
474:
469:
462:
460:
458:
454:
447:
443:
442:
438:
436:
433:
431:
428:
426:
423:
422:
418:
416:
414:
411:
406:
402:
394:
392:
388:
385:
384:
378:
375:
374:
373:Gallus gallus
368:
366:
365:
359:
357:
353:
348:
346:
345:
340:
339:
332:
330:
329:
323:
321:
317:
313:
308:
306:
305:
299:
292:
290:
288:
287:
278:
276:
272:
268:
265:
256:
254:
247:
242:
240:
238:
232:
230:
222:
218:
214:
210:
206:
202:
197:
193:
186:
184:
176:
172:
169:
165:
161:
156:
153:
148:
145:
137:
133:
128:
124:
121:
117:
116:Recombination
113:
108:
104:
101:
97:
93:
89:
85:
81:
73:
71:
67:
65:
64:
59:
58:
52:
47:
45:
41:
37:
33:
26:
21:
2389:
2384:
2374:
2334:(4): 906–9.
2331:
2325:
2319:
2284:
2279:
2269:
2244:
2240:
2234:
2199:
2194:
2184:
2149:
2144:
2134:
2126:
2121:
2086:
2081:
2071:
2036:
2031:
2021:
1986:
1981:
1971:
1936:
1931:
1921:
1886:
1881:
1871:
1836:
1831:
1783:
1778:
1728:
1723:
1677:
1673:
1629:
1623:
1613:
1568:
1563:
1505:
1501:
1491:
1470:cite journal
1437:
1396:
1390:
1384:
1331:
1327:
1317:
1276:
1272:
1262:
1227:
1223:
1213:
1160:
1156:
1146:
1136:, retrieved
1114:
1104:
1061:
1057:
1047:
1005:
998:
955:
951:
941:
906:
902:
892:
833:
829:
823:
815:
762:
758:
712:
708:
698:
655:
651:
641:
590:
586:
576:
543:
538:
476:
471:
439:
412:
407:
404:
389:
381:
379:
371:
369:
362:
360:
355:
349:
342:
336:
333:
326:
324:
319:
315:
309:
302:
300:
296:
284:
282:
273:
269:
260:
251:
233:
225:
205:Mitral cells
190:
181:
160:polymorphism
157:
149:
141:
109:
105:
80:heterozygous
77:
68:
61:
55:
48:
40:heterozygous
30:
1725:PLoS Genet.
1625:J R Soc Med
859:10871/33429
715:: 180–184.
593:(1): 4400.
356:Salmo salar
304:Salmo salar
279:In primates
152:mate choice
2430:Categories
1334:(1): 122.
1138:2022-07-28
765:(1): 107.
448:References
399:See also:
229:pheromones
213:epithelium
132:harvestman
88:homozygous
74:Hypotheses
2366:198152712
2327:Evolution
1524:1010-061X
1462:243521297
1358:1471-2148
1187:0027-8424
1078:0019-9567
1033:cite book
1025:777401230
972:1943-2631
925:0022-1503
868:0962-1083
789:1471-2148
729:1567-1348
672:1537-1719
615:2045-2322
546:: 442–6.
435:Pheromone
425:Body odor
264:Hutterite
248:In humans
227:known as
211:4: Nasal
96:pathogens
84:parasites
2416:19864282
2358:15154566
2311:19129124
2261:42727644
2226:18854296
2176:15870038
2113:17785270
2063:14667398
2013:15058398
1963:18364312
1913:18089539
1810:18700206
1757:18787687
1702:40332494
1694:19054623
1656:17541097
1540:28094212
1532:14635837
1429:19726112
1376:23768177
1309:13068773
1301:19226318
1254:18926972
1096:12874366
990:17603099
952:Genetics
933:29893971
884:51711551
876:30040159
824:Borrelia
807:31113360
737:24910107
690:35574644
633:32157133
568:10004939
560:19686174
513:15755811
419:See also
120:mutation
100:immunity
2407:2842720
2302:2679071
2217:2592554
2167:1602051
2104:2288684
2054:1809963
2004:1691569
1954:2602703
1904:2596809
1863:9364787
1854:1688704
1801:2605820
1748:2519788
1647:1885393
1605:9990056
1573:Bibcode
1421:9831558
1401:Bibcode
1392:Science
1367:3691641
1336:Bibcode
1281:Bibcode
1245:2666699
1205:1438295
1165:Bibcode
981:1950649
838:Bibcode
798:6529992
767:Bibcode
681:9132207
624:7064506
595:Bibcode
481:Bibcode
310:Female
112:alleles
42:at MHC
36:allelic
2414:
2404:
2364:
2356:
2309:
2299:
2259:
2224:
2214:
2174:
2164:
2111:
2101:
2061:
2051:
2011:
2001:
1961:
1951:
1911:
1901:
1861:
1851:
1808:
1798:
1755:
1745:
1700:
1692:
1654:
1644:
1603:
1593:
1538:
1530:
1522:
1460:
1427:
1419:
1374:
1364:
1356:
1307:
1299:
1252:
1242:
1203:
1193:
1185:
1129:
1094:
1087:166009
1084:
1076:
1023:
1013:
988:
978:
970:
931:
923:
882:
874:
866:
805:
795:
787:
735:
727:
688:
678:
670:
631:
621:
613:
566:
558:
511:
504:555479
501:
2362:S2CID
2257:S2CID
1698:S2CID
1596:15502
1536:S2CID
1458:S2CID
1425:S2CID
1305:S2CID
1196:50449
880:S2CID
564:S2CID
2412:PMID
2354:PMID
2307:PMID
2222:PMID
2172:PMID
2109:PMID
2059:PMID
2009:PMID
1959:PMID
1909:PMID
1859:PMID
1806:PMID
1753:PMID
1690:PMID
1652:PMID
1601:PMID
1528:PMID
1520:ISSN
1483:help
1417:PMID
1372:PMID
1354:ISSN
1297:PMID
1250:PMID
1201:PMID
1183:ISSN
1127:ISBN
1092:PMID
1074:ISSN
1039:link
1021:OCLC
1011:ISBN
986:PMID
968:ISSN
929:PMID
921:ISSN
872:PMID
864:ISSN
803:PMID
785:ISSN
733:PMID
725:ISSN
686:PMID
668:ISSN
629:PMID
611:ISSN
556:PMID
544:1170
509:PMID
209:Bone
142:The
138:pest
136:mite
118:and
44:loci
2402:PMC
2394:doi
2390:277
2344:hdl
2336:doi
2297:PMC
2289:doi
2285:276
2249:doi
2212:PMC
2204:doi
2200:276
2162:PMC
2154:doi
2150:272
2099:PMC
2091:doi
2087:274
2049:PMC
2041:doi
2037:270
1999:PMC
1991:doi
1987:271
1949:PMC
1941:doi
1937:275
1899:PMC
1891:doi
1887:275
1849:PMC
1841:doi
1837:264
1796:PMC
1788:doi
1784:275
1743:PMC
1733:doi
1682:doi
1642:PMC
1634:doi
1630:100
1591:PMC
1581:doi
1510:doi
1448:doi
1409:doi
1397:282
1362:PMC
1344:doi
1289:doi
1240:PMC
1232:doi
1228:364
1191:PMC
1173:doi
1119:doi
1082:PMC
1066:doi
976:PMC
960:doi
956:176
911:doi
907:109
854:hdl
846:doi
793:PMC
775:doi
717:doi
676:PMC
660:doi
619:PMC
603:doi
548:doi
499:PMC
489:doi
477:102
408:In
219:6:
215:5:
207:3:
203:2:
2432::
2410:.
2400:.
2383:.
2360:.
2352:.
2342:.
2332:58
2330:.
2305:.
2295:.
2278:.
2255:.
2245:57
2243:.
2220:.
2210:.
2193:.
2170:.
2160:.
2143:.
2107:.
2097:.
2080:.
2057:.
2047:.
2030:.
2007:.
1997:.
1980:.
1957:.
1947:.
1930:.
1907:.
1897:.
1880:.
1857:.
1847:.
1830:.
1818:^
1804:.
1794:.
1777:.
1765:^
1751:.
1741:.
1722:.
1710:^
1696:.
1688:.
1678:34
1676:.
1664:^
1650:.
1640:.
1628:.
1622:.
1599:.
1589:.
1579:.
1569:96
1562:.
1548:^
1534:.
1526:.
1518:.
1506:16
1504:.
1500:.
1474::
1472:}}
1468:{{
1456:.
1446:.
1423:.
1415:.
1407:.
1395:.
1370:.
1360:.
1352:.
1342:.
1332:13
1330:.
1326:.
1303:.
1295:.
1287:.
1277:18
1275:.
1271:.
1248:.
1238:.
1226:.
1222:.
1199:.
1189:.
1181:.
1171:.
1161:89
1159:.
1155:.
1125:,
1113:,
1090:.
1080:.
1072:.
1062:71
1060:.
1056:.
1035:}}
1031:{{
1019:.
984:.
974:.
966:.
954:.
950:.
927:.
919:.
905:.
901:.
878:.
870:.
862:.
852:.
844:.
834:27
832:.
828:.
801:.
791:.
783:.
773:.
763:19
761:.
757:.
745:^
731:.
723:.
713:26
711:.
707:.
684:.
674:.
666:.
656:39
654:.
650:.
627:.
617:.
609:.
601:.
591:10
589:.
585:.
562:.
554:.
521:^
507:.
497:.
487:.
470:.
456:^
354:,
314:,
66:.
2418:.
2396::
2368:.
2346::
2338::
2313:.
2291::
2263:.
2251::
2228:.
2206::
2178:.
2156::
2115:.
2093::
2065:.
2043::
2015:.
1993::
1965:.
1943::
1915:.
1893::
1865:.
1843::
1812:.
1790::
1759:.
1735::
1729:4
1704:.
1684::
1658:.
1636::
1607:.
1583::
1575::
1542:.
1512::
1485:)
1481:(
1464:.
1450::
1431:.
1411::
1403::
1378:.
1346::
1338::
1311:.
1291::
1283::
1256:.
1234::
1207:.
1175::
1167::
1121::
1098:.
1068::
1041:)
1027:.
992:.
962::
935:.
913::
886:.
856::
848::
840::
809:.
777::
769::
739:.
719::
692:.
662::
635:.
605::
597::
570:.
550::
515:.
491::
483::
27:.
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.