421:(ITD). Due to differing lengths and a finite conduction speed within the axons of the delay lines, different coincidence detector neurons will fire when sound comes from different positions along the azimuth. Jeffress' model proposes that two signals even from an asynchronous arrival of sound in the cochlea of each ear will converge synchronously on a coincidence detector in the auditory cortex based on the magnitude of the ITD (Fig. 2). Therefore, the ITD should correspond to an anatomical map that can be found within the brain.
1632:
314:
125:
29:
375:
receive inputs mainly from nearby cells in the same layer as the receiving cell, and also from distant connections which are fed through Layer 1. The dendrites which receive these inputs are quite distant from the cell body, and therefore they exhibit different electrical and signal-processing behaviour compared with the proximal (or feedforward) dendrites described above.
1644:
300:
potential threshold. Hence, the function of coincidence detection is to reduce the jitter caused by spontaneous neuronal activity, and while random sub-threshold stimulations from cells may not often fire coincidentally, coincident synaptic inputs derived from a unitary external stimulus ensure that a target neuron will fire as a result of the stimulus.
70:
434:
enter the nucleus laminaris dorsally while the contralateral axons enter ventrally, sounds from various positions along the azimuth correspond directly to stimulation of different depths of the nucleus laminaris. From this information, a neural map of auditory space was formed. The function of the nucleus laminaris parallels that of the
383:
received within a short period of time (i.e. before the overall voltage decays to background), the voltage of the segment will rise above a threshold, giving rise to a non-linear dendritic spike, which travels, effectively undiminished, all the way to the cell body, and which causes it to become partially depolarised.
497:. As a result, both synapses strengthen. The prolonged depolarization needed for the expulsion of Mg from NMDA receptors requires a high frequency stimulation. Associativity becomes a factor because this can be achieved through two simultaneous inputs that may not be strong enough to activate LTP by themselves.
500:
Besides the NMDA-receptor based processes, further cellular mechanisms allow of the association between two different input signals converging on the same neuron, in a defined timeframe. Upon a simultaneous increase in the intracellular concentrations of cAMP and Ca, a transcriptional coactivator
433:
of the ears travels to the ipsilateral nucleus magnocellularis. From here, the signals project ipsilaterally and contralaterally to two nucleus laminari. Each nucleus laminaris contains coincidence detectors that receive auditory input from the left and the right ear. Since the ipsilateral axons
374:
The above description applies well to feedforward inputs to neurons, which provide inputs from either sensory nerves or lower-level regions in the brain. About 90% of interneural connections are, however, not feedforward but predictive (or modulatory, or attentional) in nature. These connections
382:
long) of distal dendrite, the reaction to activations coming in on synapses to the dendritic spines acts to raise the overall local potential with each incoming signal. This rising potential acts against a background of decay in the potential back to the resting level. If sufficient signals are
299:
of a target neuron over the threshold required to create an action potential. Conversely, if the two inputs temporally arrive too far apart, the depolarization of the first input may have time to drop significantly, preventing the membrane potential of the target neuron from reaching the action
416:
proposed that some organisms may have a collection of neurons that receive auditory input from each ear. The neural pathways to these neurons are called delay lines. Jeffress claimed that the neurons that the delay lines link act as coincidence detectors by firing maximally when receiving
286:
Coincidence detection relies on separate inputs converging on a common target. For example (Fig. 1), in a basic neural circuit with two input neurons—A and B—that have excitatory synaptic terminals converging on a single output neuron (C), if each input neuron's
275:
386:
This is perhaps the most important form of dendritic coincidence detection in the brain. The more easily understood proximal activation acts over much longer time periods, and is thus much less sensitive to the time factor in coincidence detection.
399:
Fig. 2: If a sound arrives at the left ear before the right ear, the impulse in the left auditory tract will reach X sooner than the impulse in the right auditory tract reaches Y. Neurons 4 or 5 may therefore receive coincident
250:
encodes information by detecting the occurrence of temporally close but spatially distributed input signals. Coincidence detectors influence neuronal information processing by reducing temporal
335:
142:
42:
450:
postulated that synaptic efficiency will increase through repeated and persistent stimulation of a postsynaptic cell by a presynaptic cell. This is often informally summarized as "
88:
458:. Studies of LTP on multiple presynaptic cells stimulating a postsynaptic cell uncovered the property of associativity. A weak neuronal stimulation onto a
48:
1296:
189:
1336:
161:
548:. Glutamate released from the parallel fibers activates AMPA receptors which depolarize the postsynaptic cell. The parallel fibers also activate
168:
295:
at C, then C cannot fire unless the two inputs from A and B are temporally close. The synchronous arrival of these two inputs may push the
417:
simultaneous inputs from both ears. When a sound is heard, sound waves may reach the ears at different times. This is referred to as the
462:
may not induce long-term potentiation. However, this same stimulation paired with a simultaneous strong stimulation from another neuron
396:
175:
1381:
950:
Willoughby, Debbie; Cooper, Dermot M. F. (July 2007). "Organization and Ca2+ regulation of adenylyl cyclases in cAMP microdomains".
875:
361:
288:
226:
208:
106:
56:
157:
586:
466:. This process suggests that two neuronal pathways converging on the same cell may both strengthen if stimulated coincidentally.
536:
Long-term depression also works through associative properties although it is not always the reverse process of LTP. LTD in the
1670:
1580:
339:
146:
794:
Zupanc, G.K.H. 2004. Behavioral
Neurobiology: An Integrative Approach. Oxford University Press: Oxford, UK. pp. 133-150
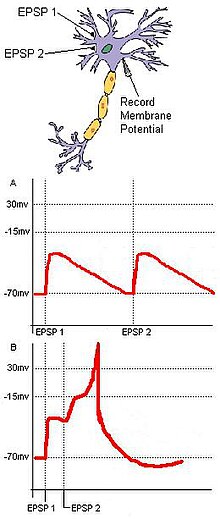
282:. Two EPSPs innervated in rapid succession sum to produce a larger EPSP, or an action potential in the postsynaptic cell.
1329:
568:
279:
324:
1297:
https://web.archive.org/web/20040519194818/http://bbsonline.cup.cam.ac.uk/Preprints/OldArchive/bbs.neur4.crepel.html
1585:
1570:
893:"TORC1 is a calcium- and cAMP-sensitive coincidence detector involved in hippocampal long-term synaptic plasticity"
418:
182:
343:
328:
135:
1386:
1005:"The role of Ca2+/calmodulin-stimulable adenylyl cyclases as molecular coincidence detectors in memory formation"
891:
Kovacs, K. A.; Steullet, P.; Steinmann, M.; Do, K. Q.; Magistretti, P. J.; Halfon, O.; Cardinaux, J. -R. (2007).
485:. The removal of the Mg block allows the flow of Ca into the cell. A large elevation of calcium levels activate
1648:
1590:
1443:
1111:
Neve, Kim A.; Seamans, Jeremy K.; Trantham-Davidson, Heather (August 2004). "Dopamine receptor signaling".
254:
and spontaneous activity, allowing the creation of variable associations between separate neural events in
1636:
1600:
1359:
1322:
1120:
959:
601:
556:
and DAG. The climbing fibers stimulate a large increase in postsynaptic Ca levels when activated. The Ca,
474:
455:
1416:
557:
435:
1493:
1426:
1406:
1401:
904:
813:
749:
655:
596:
1308:
1125:
964:
1560:
1498:
1483:
1448:
581:
17:
571:
to internalize AMPA receptors and decrease the sensitivity of the postsynaptic cell to glutamate.
1523:
1453:
1421:
1396:
1154:
1050:
845:
804:
Frey, Uwe; Morris, Richard G. M. (February 1997). "Synaptic tagging and long-term potentiation".
621:
405:
296:
1544:
1285:
1250:
1207:
1146:
1138:
1093:
1085:
1042:
1024:
985:
977:
932:
871:
837:
829:
777:
718:
683:
510:
506:
413:
1610:
1605:
1518:
1275:
1240:
1232:
1197:
1189:
1130:
1077:
1032:
1016:
969:
922:
912:
821:
767:
757:
710:
673:
663:
463:
459:
422:
292:
1575:
1478:
1473:
644:"On the relationship between synaptic input and spike output jitter in individual neurons"
545:
541:
513:
activation, might also account for the detection of the repetitive stimulation of a given
486:
1178:"Two Coincidence Detectors for Spike Timing-Dependent Plasticity in Somatosensory Cortex"
1037:
1004:
908:
817:
753:
659:
1595:
1468:
1463:
1411:
1245:
1221:"Long-term potentiation of GABAergic synaptic transmission in neonatal rat hippocampus"
1220:
1202:
1177:
927:
892:
591:
509:
into long term changes such as LTP. This cellular mechanism, through calcium-dependent
247:
1280:
1263:
1081:
772:
737:
1664:
1458:
1391:
1345:
1236:
678:
643:
611:
490:
482:
1158:
1054:
1615:
1565:
1503:
1193:
1068:
Hanoune, J.; Defer, N. (2001). "Regulation and role of adenylyl cyclase isoforms".
849:
616:
606:
549:
263:
259:
1539:
1508:
1488:
564:
481:
requires a prolonged depolarization that can expel the Mg block of postsynaptic
478:
447:
313:
274:
124:
973:
742:
Proceedings of the
National Academy of Sciences of the United States of America
648:
Proceedings of the
National Academy of Sciences of the United States of America
1513:
537:
430:
395:
1142:
1089:
1028:
981:
833:
917:
762:
494:
426:
1254:
1211:
1150:
1097:
1046:
989:
936:
722:
1289:
1134:
1020:
841:
781:
687:
668:
1376:
1264:"Coincidence detection in the auditory system: 50 years after Jeffress"
514:
505:) becomes activated, that converts the temporal coincidence of the two
409:
1370:
1176:
Bender, V. A.; Bender, K. J.; Brasier, D. J.; Feldman, D. E. (2006).
825:
714:
528:) has been implicated in memory formation as a coincidence detector.
255:
251:
243:
870:(3 ed.). Sunderland, MA: Sinauer Associates. pp. 575–608.
502:
394:
379:
273:
1314:
738:"Axonal delay lines for time measurement in the owl's brainstem"
701:
Jeffress, L. A. (1948). "A place theory of sound localization".
1318:
307:
118:
63:
22:
493:. This increases the sensitivity of the postsynaptic cell to
404:
Coincidence detection has been shown to be a major factor in
1309:
552:
glutamate receptors that release the second messengers IP
258:. The study of coincidence detectors has been crucial in
454:". The theory was validated in part by the discovery of
84:
429:
shows that this is true. Sensory information from the
1113:
Journal of
Receptor and Signal Transduction Research
1003:
Mons, N.; Guillou, J.-L.; Jaffard, R. (1999-04-01).
489:
that ultimately increase the number of postsynaptic
1553:
1532:
1436:
1352:
703:
149:. Unsourced material may be challenged and removed.
79:
may be too technical for most readers to understand
1219:Caillard, O.; Ben-Ari, Y.; Gaiarsa, J. L. (1999).
1262:Joris, P. X.; Smith, P. H.; Yin, T. C. (1998).
897:Proceedings of the National Academy of Sciences
262:with regards to understanding the formation of
1330:
861:
859:
642:Marsálek, P.; Koch, C.; Maunsell, J. (1997).
470:Molecular mechanism of long-term potentiation
8:
1070:Annual Review of Pharmacology and Toxicology
532:Molecular mechanism of long-term depression
342:. Unsourced material may be challenged and
57:Learn how and when to remove these messages
1337:
1323:
1315:
1279:
1244:
1201:
1124:
1036:
963:
926:
916:
771:
761:
677:
667:
520:Adenylyl cyclase (also commonly known as
362:Learn how and when to remove this message
227:Learn how and when to remove this message
209:Learn how and when to remove this message
107:Learn how and when to remove this message
91:, without removing the technical details.
634:
452:cells that fire together, wire together
158:"Coincidence detection in neurobiology"
1311:- Online interactive tutorial (Flash)
540:requires a coincident stimulation of
442:Synaptic plasticity and associativity
412:plane in several organisms. In 1948,
89:make it understandable to non-experts
7:
1643:
1009:Cellular and Molecular Life Sciences
378:In a short section (perhaps 40
340:adding citations to reliable sources
147:adding citations to reliable sources
270:Principles of coincidence detection
14:
1082:10.1146/annurev.pharmtox.41.1.145
736:Carr, C. E.; Konishi, M. (1988).
38:This article has multiple issues.
1642:
1631:
1630:
1237:10.1111/j.1469-7793.1999.0109r.x
587:Earth Coincidence Control Office
312:
123:
68:
27:
1387:Central pattern generator (CPG)
134:needs additional citations for
46:or discuss these issues on the
16:For the electronic device, see
1581:Frog hearing and communication
1194:10.1523/JNEUROSCI.0176-06.2006
280:Spatial and temporal summation
1:
1281:10.1016/S0896-6273(00)80643-1
464:will strengthen both synapses
304:Distal coincidence detection
569:signal transduction pathway
1687:
1586:Infrared sensing in snakes
1571:Jamming avoidance response
974:10.1152/physrev.00049.2006
419:interaural time difference
15:
1626:
1225:The Journal of Physiology
1591:Caridoid escape reaction
291:is sub-threshold for an
1444:Theodore Holmes Bullock
1182:Journal of Neuroscience
918:10.1073/pnas.0607524104
763:10.1073/pnas.85.21.8311
1671:Neuroethology concepts
1601:Surface wave detection
602:Long-term potentiation
456:long-term potentiation
401:
283:
1417:Anti-Hebbian learning
1135:10.1081/RRS-200029981
1021:10.1007/s000180050311
952:Physiological Reviews
866:Purves, Dale (2004).
669:10.1073/pnas.94.2.735
436:medial superior olive
398:
277:
240:Coincidence detection
1494:Bernhard Hassenstein
1427:Ultrasound avoidance
1402:Fixed action pattern
1365:Coincidence detector
597:Long-term depression
336:improve this section
143:improve this article
1561:Animal echolocation
1499:Werner E. Reichardt
1449:Walter Heiligenberg
909:2007PNAS..104.4700K
818:1997Natur.385..533F
754:1988PNAS...85.8311C
660:1997PNAS...94..735M
582:Coincidence circuit
567:work together in a
246:process in which a
18:Coincidence circuit
1524:Fernando Nottebohm
1422:Sound localization
1397:Lateral inhibition
622:Sound localization
423:Masakazu Konishi's
406:sound localization
402:
391:Sound localization
297:membrane potential
284:
264:computational maps
1658:
1657:
1545:Slice preparation
1407:Krogh's Principle
1382:Feature detection
1231:(Pt 1): 109–119.
1188:(16): 4166–4177.
903:(11): 4700–4705.
812:(6616): 533–536.
748:(21): 8311–8315.
526:adenylate cyclase
511:adenylate cyclase
507:second messengers
414:Lloyd A. Jeffress
372:
371:
364:
237:
236:
229:
219:
218:
211:
193:
117:
116:
109:
61:
1678:
1646:
1645:
1634:
1633:
1611:Mechanoreception
1606:Electroreception
1519:Masakazu Konishi
1484:Jörg-Peter Ewert
1339:
1332:
1325:
1316:
1293:
1283:
1274:(6): 1235–1238.
1258:
1248:
1215:
1205:
1163:
1162:
1128:
1108:
1102:
1101:
1065:
1059:
1058:
1040:
1000:
994:
993:
967:
947:
941:
940:
930:
920:
888:
882:
881:
863:
854:
853:
826:10.1038/385533a0
801:
795:
792:
786:
785:
775:
765:
733:
727:
726:
715:10.1037/h0061495
698:
692:
691:
681:
671:
639:
460:pyramidal neuron
367:
360:
356:
353:
347:
316:
308:
293:action potential
232:
225:
214:
207:
203:
200:
194:
192:
151:
127:
119:
112:
105:
101:
98:
92:
72:
71:
64:
53:
31:
30:
23:
1686:
1685:
1681:
1680:
1679:
1677:
1676:
1675:
1661:
1660:
1659:
1654:
1622:
1576:Vision in toads
1549:
1528:
1479:Erich von Holst
1474:Karl von Frisch
1432:
1348:
1343:
1305:
1261:
1218:
1175:
1172:
1170:Further reading
1167:
1166:
1126:10.1.1.465.5011
1110:
1109:
1105:
1067:
1066:
1062:
1002:
1001:
997:
965:10.1.1.336.3746
958:(3): 965–1010.
949:
948:
944:
890:
889:
885:
878:
865:
864:
857:
803:
802:
798:
793:
789:
735:
734:
730:
700:
699:
695:
641:
640:
636:
631:
626:
577:
561:
555:
546:climbing fibers
542:parallel fibers
534:
487:protein kinases
472:
444:
393:
368:
357:
351:
348:
333:
317:
306:
272:
233:
222:
221:
220:
215:
204:
198:
195:
152:
150:
140:
128:
113:
102:
96:
93:
85:help improve it
82:
73:
69:
32:
28:
21:
12:
11:
5:
1684:
1682:
1674:
1673:
1663:
1662:
1656:
1655:
1653:
1652:
1640:
1627:
1624:
1623:
1621:
1620:
1619:
1618:
1608:
1603:
1598:
1596:Vocal learning
1593:
1588:
1583:
1578:
1573:
1568:
1563:
1557:
1555:
1551:
1550:
1548:
1547:
1542:
1536:
1534:
1530:
1529:
1527:
1526:
1521:
1516:
1511:
1506:
1501:
1496:
1491:
1486:
1481:
1476:
1471:
1469:Donald Kennedy
1466:
1464:Donald Griffin
1461:
1456:
1454:Niko Tinbergen
1451:
1446:
1440:
1438:
1434:
1433:
1431:
1430:
1424:
1419:
1414:
1412:Hebbian theory
1409:
1404:
1399:
1394:
1389:
1384:
1379:
1374:
1367:
1362:
1356:
1354:
1350:
1349:
1344:
1342:
1341:
1334:
1327:
1319:
1313:
1312:
1304:
1303:External links
1301:
1300:
1299:
1294:
1259:
1216:
1171:
1168:
1165:
1164:
1119:(3): 165–205.
1103:
1060:
1015:(4): 525–533.
995:
942:
883:
876:
855:
796:
787:
728:
693:
654:(2): 735–740.
633:
632:
630:
627:
625:
624:
619:
614:
609:
604:
599:
594:
592:Hebbian theory
589:
584:
578:
576:
573:
559:
553:
533:
530:
522:adenyl cyclase
501:called TORC1 (
491:AMPA receptors
483:NMDA receptors
471:
468:
443:
440:
392:
389:
370:
369:
320:
318:
311:
305:
302:
271:
268:
266:in the brain.
248:neural circuit
235:
234:
217:
216:
131:
129:
122:
115:
114:
76:
74:
67:
62:
36:
35:
33:
26:
13:
10:
9:
6:
4:
3:
2:
1683:
1672:
1669:
1668:
1666:
1651:
1650:
1641:
1639:
1638:
1629:
1628:
1625:
1617:
1614:
1613:
1612:
1609:
1607:
1604:
1602:
1599:
1597:
1594:
1592:
1589:
1587:
1584:
1582:
1579:
1577:
1574:
1572:
1569:
1567:
1564:
1562:
1559:
1558:
1556:
1552:
1546:
1543:
1541:
1538:
1537:
1535:
1531:
1525:
1522:
1520:
1517:
1515:
1512:
1510:
1507:
1505:
1502:
1500:
1497:
1495:
1492:
1490:
1487:
1485:
1482:
1480:
1477:
1475:
1472:
1470:
1467:
1465:
1462:
1460:
1459:Konrad Lorenz
1457:
1455:
1452:
1450:
1447:
1445:
1442:
1441:
1439:
1435:
1428:
1425:
1423:
1420:
1418:
1415:
1413:
1410:
1408:
1405:
1403:
1400:
1398:
1395:
1393:
1392:NMDA receptor
1390:
1388:
1385:
1383:
1380:
1378:
1375:
1373:
1372:
1368:
1366:
1363:
1361:
1358:
1357:
1355:
1351:
1347:
1346:Neuroethology
1340:
1335:
1333:
1328:
1326:
1321:
1320:
1317:
1310:
1307:
1306:
1302:
1298:
1295:
1291:
1287:
1282:
1277:
1273:
1269:
1265:
1260:
1256:
1252:
1247:
1242:
1238:
1234:
1230:
1226:
1222:
1217:
1213:
1209:
1204:
1199:
1195:
1191:
1187:
1183:
1179:
1174:
1173:
1169:
1160:
1156:
1152:
1148:
1144:
1140:
1136:
1132:
1127:
1122:
1118:
1114:
1107:
1104:
1099:
1095:
1091:
1087:
1083:
1079:
1075:
1071:
1064:
1061:
1056:
1052:
1048:
1044:
1039:
1034:
1030:
1026:
1022:
1018:
1014:
1010:
1006:
999:
996:
991:
987:
983:
979:
975:
971:
966:
961:
957:
953:
946:
943:
938:
934:
929:
924:
919:
914:
910:
906:
902:
898:
894:
887:
884:
879:
877:9780878937257
873:
869:
862:
860:
856:
851:
847:
843:
839:
835:
831:
827:
823:
819:
815:
811:
807:
800:
797:
791:
788:
783:
779:
774:
769:
764:
759:
755:
751:
747:
743:
739:
732:
729:
724:
720:
716:
712:
708:
704:
697:
694:
689:
685:
680:
675:
670:
665:
661:
657:
653:
649:
645:
638:
635:
628:
623:
620:
618:
615:
613:
612:Neuroethology
610:
608:
605:
603:
600:
598:
595:
593:
590:
588:
585:
583:
580:
579:
574:
572:
570:
566:
562:
551:
547:
543:
539:
531:
529:
527:
523:
518:
516:
512:
508:
504:
498:
496:
492:
488:
484:
480:
476:
469:
467:
465:
461:
457:
453:
449:
441:
439:
437:
432:
428:
424:
420:
415:
411:
407:
397:
390:
388:
384:
381:
376:
366:
363:
355:
352:November 2013
345:
341:
337:
331:
330:
326:
321:This section
319:
315:
310:
309:
303:
301:
298:
294:
290:
281:
276:
269:
267:
265:
261:
257:
253:
249:
245:
241:
231:
228:
213:
210:
202:
199:November 2013
191:
188:
184:
181:
177:
174:
170:
167:
163:
160: –
159:
155:
154:Find sources:
148:
144:
138:
137:
132:This article
130:
126:
121:
120:
111:
108:
100:
97:November 2013
90:
86:
80:
77:This article
75:
66:
65:
60:
58:
51:
50:
45:
44:
39:
34:
25:
24:
19:
1647:
1635:
1616:Lateral line
1566:Waggle dance
1504:Eric Knudsen
1369:
1364:
1271:
1267:
1228:
1224:
1185:
1181:
1116:
1112:
1106:
1073:
1069:
1063:
1012:
1008:
998:
955:
951:
945:
900:
896:
886:
868:Neuroscience
867:
809:
805:
799:
790:
745:
741:
731:
709:(1): 35–39.
706:
702:
696:
651:
647:
637:
617:Quantum mind
607:Neurobiology
550:metabotropic
535:
525:
521:
519:
499:
473:
451:
445:
438:in mammals.
403:
385:
377:
373:
358:
349:
334:Please help
322:
285:
260:neuroscience
239:
238:
223:
205:
196:
186:
179:
172:
165:
153:
141:Please help
136:verification
133:
103:
94:
78:
54:
47:
41:
40:Please help
37:
1540:Patch clamp
1509:Eric Kandel
1489:Franz Huber
1360:Feedforward
1076:: 145–174.
479:hippocampus
448:Donald Hebb
1514:Nobuo Suga
1429:in insects
629:References
538:cerebellum
431:hair cells
408:along the
169:newspapers
43:improve it
1143:1079-9893
1121:CiteSeerX
1090:0362-1642
1029:1420-682X
982:0031-9333
960:CiteSeerX
834:0028-0836
495:glutamate
446:In 1949,
427:barn owls
425:study on
323:does not
49:talk page
1665:Category
1637:Category
1377:Instinct
1353:Concepts
1255:10373693
1212:16624937
1159:12407397
1151:15521361
1098:11264454
1055:10849274
1047:10357223
1038:11147090
990:17615394
937:17360587
723:18904764
575:See also
278:Fig. 1:
244:neuronal
1649:Commons
1554:Systems
1533:Methods
1290:9883717
1246:2269393
1203:3071735
928:1838663
905:Bibcode
850:4339789
842:9020359
814:Bibcode
782:3186725
750:Bibcode
688:9012854
656:Bibcode
515:synapse
477:in the
410:azimuth
400:inputs.
344:removed
329:sources
183:scholar
83:Please
1437:People
1371:Umwelt
1288:
1268:Neuron
1253:
1243:
1210:
1200:
1157:
1149:
1141:
1123:
1096:
1088:
1053:
1045:
1035:
1027:
988:
980:
962:
935:
925:
874:
848:
840:
832:
806:Nature
780:
773:282419
770:
721:
686:
676:
563:, and
256:memory
252:jitter
185:
178:
171:
164:
156:
1155:S2CID
1051:S2CID
846:S2CID
679:19583
503:CRTC1
242:is a
190:JSTOR
176:books
1286:PMID
1251:PMID
1208:PMID
1147:PMID
1139:ISSN
1094:PMID
1086:ISSN
1043:PMID
1025:ISSN
986:PMID
978:ISSN
933:PMID
872:ISBN
838:PMID
830:ISSN
778:PMID
719:PMID
684:PMID
544:and
524:and
327:any
325:cite
289:EPSP
162:news
1276:doi
1241:PMC
1233:doi
1229:518
1198:PMC
1190:doi
1131:doi
1078:doi
1033:PMC
1017:doi
970:doi
923:PMC
913:doi
901:104
822:doi
810:385
768:PMC
758:doi
711:doi
674:PMC
664:doi
565:DAG
475:LTP
338:by
145:by
87:to
1667::
1284:.
1272:21
1270:.
1266:.
1249:.
1239:.
1227:.
1223:.
1206:.
1196:.
1186:26
1184:.
1180:.
1153:.
1145:.
1137:.
1129:.
1117:24
1115:.
1092:.
1084:.
1074:41
1072:.
1049:.
1041:.
1031:.
1023:.
1013:55
1011:.
1007:.
984:.
976:.
968:.
956:87
954:.
931:.
921:.
911:.
899:.
895:.
858:^
844:.
836:.
828:.
820:.
808:.
776:.
766:.
756:.
746:85
744:.
740:.
717:.
707:41
705:.
682:.
672:.
662:.
652:94
650:.
646:.
558:IP
517:.
380:μm
52:.
1338:e
1331:t
1324:v
1292:.
1278::
1257:.
1235::
1214:.
1192::
1161:.
1133::
1100:.
1080::
1057:.
1019::
992:.
972::
939:.
915::
907::
880:.
852:.
824::
816::
784:.
760::
752::
725:.
713::
690:.
666::
658::
560:3
554:3
365:)
359:(
354:)
350:(
346:.
332:.
230:)
224:(
212:)
206:(
201:)
197:(
187:·
180:·
173:·
166:·
139:.
110:)
104:(
99:)
95:(
81:.
59:)
55:(
20:.
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.