182:(1992), described neurons in the entorhinal cortex as having relatively large and fuzzy place fields. But the Mosers thought it was possible that a different result would be obtained if recordings were made from a different part of the entorhinal cortex. The entorhinal cortex is a strip of tissue running along the back edge of the rat brain from the ventral to the dorsal sides. Anatomical studies had shown that different sectors of the entorhinal cortex project to different levels of the hippocampus: the dorsal end of the EC projects to the dorsal hippocampus, the ventral end to the ventral hippocampus. This was relevant because several studies had shown that place cells in the dorsal hippocampus have considerably sharper place fields than cells from more ventral levels. But every study of entorhinal spatial activity before 2004 had made use of electrodes implanted near the ventral end of the EC. Accordingly, together with Marianne Fyhn, Sturla Molden, and Menno Witter, the Mosers set out to examine spatial firing from the different dorsal-to-ventral levels of the entorhinal cortex. They found that in the dorsal part of medial entorhinal cortex (MEC), cells had sharply defined place fields like in the hippocampus but the cells fired at multiple locations. The arrangement of the firing fields showed hints of regularity, but the size of the environment was too small for spatial periodicity to be visible in this study.
292:. A number of theoretical models have explored mechanisms by which path integration could be performed by neural networks. In most models, such as those of Samsonovich and McNaughton (1997) or Burak and Fiete (2009), the principal ingredients are (1) an internal representation of position, (2) internal representations of the speed and direction of movement, and (3) a mechanism for shifting the encoded position by the right amount when the animal moves. Because cells in the MEC encode information about position (grid cells) and movement (head direction cells and conjunctive position-by-direction cells), this area is currently viewed as the most promising candidate for the place in the brain where path integration occurs. However, the question remains unresolved, as in humans the entorhinal cortex does not appear to be required for path integration. Burak and Fiete (2009) showed that a computational simulation of the grid cell system was capable of performing path integration to a high level of accuracy. However, more recent theoretical work has suggested that grid cells might perform a more general denoising process not necessarily related to spatial processing.
222:
typically show different grid spacings. Cells located more ventrally (farther from the dorsal border of the MEC) generally have larger firing fields at each grid vertex, and correspondingly greater spacing between the grid vertices. The total range of grid spacings is not well established: the initial report described a roughly twofold range of grid spacings (from 39 cm to 73 cm) across the dorsalmost part (upper 25%) of the MEC, but there are indications of considerably larger grid scales in more ventral zones. Brun
186:
the phase of the grid (the offset of the grid vertices relative to the x and y axes) appeared to be randomly distributed between cells. The periodic firing pattern was expressed independently of the configuration of landmarks, in darkness as well as in the presence of visible landmarks and independently of changes in the animal’s speed and direction, leading the authors to suggest that grid cells expressed a path-integration-dependent dynamic computation of the animal’s location.
303:
246:, in which many cells alter their firing rates but the majority of cells retain place fields in the same locations as before. This was examined using simultaneous recordings of hippocampal and entorhinal cells, and found that in situations where the hippocampus shows rate remapping, grid cells show unaltered firing patterns, whereas when the hippocampus shows complete remapping, grid cell firing patterns show unpredictable shifts and rotations.
226:(2008) recorded grid cells from multiple levels in rats running along an 18-meter track, and found that the grid spacing expanded from about 25 cm in their dorsalmost sites to about 3 m at the ventralmost sites. These recordings only extended 3/4 of the way to the ventral tip, so it is possible that even larger grids exist. Such multi-scale representations have been shown to be information theoretically desirable.
202:
110:
38:
230:
to rotate by the same amount. Grid patterns appear on the first entrance of an animal into a novel environment, and then usually remain stable. When an animal is moved into a completely different environment, grid cells maintain their grid spacing, and the grids of neighboring cells maintain their relative offsets.
214:(dMEC), but cells in different layers tend to differ in other respects. Layer II contains the largest density of pure grid cells, in the sense that they fire equally regardless of the direction in which an animal traverses a grid location. Grid cells from deeper layers are intermingled with conjunctive cells and
278:
composed of stellate cells. Most grid cells from layer III do not precess, but their spike activity is largely confined to half of the theta cycle. The grid cell phase precession is not derived from the hippocampus, because it continues to appear in animals whose hippocampus has been inactivated by
229:
Grid-cell activity does not require visual input, since grid patterns remain unchanged when all the lights in an environment are turned off. But when visual cues are present they exert strong control over the alignment of the grids: rotating a cue card on the wall of a cylinder causes grid patterns
209:
Grid cells are neurons that fire when a freely moving animal traverses a set of small regions (firing fields) which are roughly equal in size and arranged in a periodic triangular array that covers the entire available environment. Cells with this firing pattern have been found in all layers of the
221:
Grid cells that lie next to one another (i.e., cells recorded from the same electrode) usually show the same grid spacing and orientation, but their grid vertices are displaced from one another by apparently random offsets. But cells recorded from separate electrodes at a distance from one another
185:
The next set of experiments, reported in 2005, made use of a larger environment, which led to the recognition that the cells were actually firing in a hexagonal grid pattern. The study showed that cells at similar dorsal-to-ventral MEC levels had similar grid spacing and grid orientation, but that
242:—that is, the pattern of place fields reorganizes in a way that bears no detectable resemblance to the pattern in the original environment. If the features of an environment are altered less radically, however, the place field pattern may show a lesser degree of change, referred to as
327:
Firing fields are generally equally spaced apart, such that the distance from one firing field to all six adjacent firing fields is approximately the same (though when an environment is resized, the field spacing may shrink or expand differently in different directions; Barry et al.
129:. These marks accumulate over time to form a set of small clusters, which in turn form the vertices of a grid of equilateral triangles. The regular triangle pattern distinguishes grid cells from other types of cells that show spatial firing. By contrast, if a
57:
that fires at regular intervals as an animal navigates an open area, allowing it to understand its position in space by storing and integrating information about location, distance, and direction. Grid cells have been found in many animals, including
133:
from the rat hippocampus is examined in the same way, then the marks will frequently only form one cluster (one "place field") in a given environment, and even when multiple clusters are seen, there is no perceptible regularity in their arrangement.
101:
for their discoveries of cells that constitute a positioning system in the brain. The arrangement of spatial firing fields, all at equal distances from their neighbors, led to a hypothesis that these cells encode a neural representation of
274:—that is, their spike activity advances from late to early phases of the theta cycle as an animal passes through a grid vertex. A recent model of grid cell activity explained this phase precession by assuming the presence of 1-dimensional
90:
287:
Many species of mammals can keep track of spatial location even in the absence of visual, auditory, olfactory, or tactile cues, by integrating their movements—the ability to do this is referred to in the literature as
177:
Because the entorhinal cortex provides by far the largest input to the hippocampus, it was clearly important to understand the spatial firing properties of entorhinal neurons. The earliest studies, such as Quirk
270:, the central controller of theta. Grid cells, like hippocampal place cells, show strong theta modulation. Grid cells from layer II of the MEC also resemble hippocampal place cells in that they show
867:
Maurer, A. P.; Vanrhoads, S. R.; Sutherland, G. R.; Lipa, P.; McNaughton, B. L. (2005). "Self-motion and the origin of differential spatial scaling along the septo-temporal axis of the hippocampus".
956:
Sargolini, F.; Fyhn, M.; Hafting, T.; McNaughton, B. L.; Witter, M. P.; Moser, M. -B.; Moser, E. I. (2006). "Conjunctive
Representation of Position, Direction, and Velocity in Entorhinal Cortex".
125:
and collects recordings as the rat moves around freely in an open arena. The resulting data can be visualized by marking the rat's position on a map of the arena every time that neuron fires an
174:. This theory aroused a great deal of interest, and motivated hundreds of experimental studies aimed at clarifying the role of the hippocampus in spatial memory and spatial navigation.
158:
of the cell. This discovery, although controversial at first, led to a series of investigations that culminated in the 1978 publication of a book by O'Keefe and his colleague
350:
314:, a grid cell has multiple firing fields, with regular spacing, which tessellate the environment in a hexagonal pattern. The unique properties of grid cells are as follows:
335:
The grid cells are anchored to external landmarks, but persist in darkness, suggesting that grid cells may be part of a self-motion–based map of the spatial environment.
318:
Grid cells have firing fields dispersed over the entire environment (in contrast to place fields which are restricted to certain specific regions of the environment)
604:
Jacobs, J.; Weidemann, C. T.; Miller, J. F.; Solway, A.; Burke, J. F.; Wei, X. X.; Suthana, N.; Sperling, M. R.; Sharan, A. D.; Fried, I.; Kahana, M. J. (2013).
106:. The discovery also suggested a mechanism for dynamic computation of self-position based on continuously updated information about position and direction.
41:
Trajectory of a rat through a square environment is shown in black. Red dots indicate locations at which a particular entorhinal grid cell fired.
1238:
Hafting, T.; Fyhn, M.; Bonnevie, T.; Moser, M. B.; Moser, E. I. (2008). "Hippocampus-independent phase precession in entorhinal grid cells".
190:
94:
1136:
Fyhn, M.; Hafting, T.; Treves, A.; Moser, M. B.; Moser, E. I. (2007). "Hippocampal remapping and grid realignment in entorhinal cortex".
218:(i.e. in layers III, V and VI there are cells with a grid-like pattern that fire only when the animal is facing a particular direction).
258:, which has a frequency range of about 6–9 Hz in rats. The entorhinal cortex is no exception: like the hippocampus, it receives
837:
1469:
Sreenivasan, S; Fiete, I (2011). "Grid cells generate an analog error-correcting code for singularly precise neural computation".
396:
Hafting, T.; Fyhn, M.; Molden, S.; Moser, M. B.; Moser, E. I. (2005). "Microstructure of a spatial map in the entorhinal cortex".
746:
143:
98:
496:
Yartsev, M. M.; Witter, M. P.; Ulanovsky, N. (2011). "Grid cells without theta oscillations in the entorhinal cortex of bats".
166:(a phrase that also appeared in the title of the 1971 paper)—the book argued that the hippocampal neural network instantiates
710:
O'Keefe, D. J. (1971). "The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat".
910:
Fyhn, M.; Molden, S.; Witter, M. P.; Moser, E. I.; Moser, M. -B. (2004). "Spatial
Representation in the Entorhinal Cortex".
331:
Firing fields are equally positioned, such that the six neighboring fields are located at approximately 60 degree increments
775:"The positional firing properties of medial entorhinal neurons: description and comparison with hippocampal place cells"
1517:
154:—cells that fire action potentials when an animal passes through a specific small region of space, which is called the
238:
When a rat is moved to a different environment, the spatial activity patterns of hippocampal place cells usually show
113:
Grid cells derive their name from the fact that connecting the centers of their firing fields gives a triangular grid.
1561:
255:
779:
299:, which may make the associations between place and events that are needed for the formation of memories.
295:
Hafting et al. (2005) suggested that a place code is computed in the entorhinal cortex and fed into the
1189:"Episodic Memories: How do the Hippocampus and the Entorhinal Ring Attractors Cooperate to Create Them?"
999:
Brun, V. H.; Solstad, T.; Kjelstrup, K. B.; Fyhn, M.; Witter, M. P.; Moser, E. I.; Moser, M. B. (2008).
118:
1412:"Neural basis of the cognitive map: Path integration does not require hippocampus or entorhinal cortex"
1423:
1364:
1247:
1145:
965:
919:
666:
560:
505:
405:
215:
1089:"The effects of changes in the environment on the spatial firing of hippocampal complex-spike cells"
1546:
1551:
1494:
1354:
1271:
1169:
1069:
892:
849:
529:
478:
429:
31:
302:
1486:
1451:
1392:
1320:
1263:
1220:
1161:
1118:
1061:
1022:
981:
935:
884:
841:
806:
727:
692:
635:
586:
521:
470:
421:
322:
275:
211:
122:
54:
1541:
1478:
1441:
1431:
1382:
1372:
1310:
1302:
1255:
1210:
1200:
1153:
1108:
1100:
1053:
1012:
973:
927:
876:
833:
796:
788:
719:
682:
674:
625:
617:
576:
568:
513:
460:
413:
289:
271:
171:
126:
63:
1521:
103:
86:
1427:
1368:
1251:
1149:
969:
923:
670:
564:
509:
409:
205:
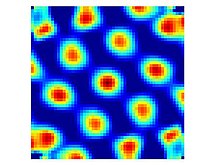
Spatial autocorrelogram of the neuronal activity of the grid cell from the first figure.
1556:
1446:
1411:
1387:
1342:
1315:
1306:
1291:"Path integration and cognitive mapping in a continuous attractor neural network model"
1290:
1215:
1188:
1113:
1104:
1088:
801:
792:
774:
687:
654:
630:
605:
581:
548:
59:
752:
201:
189:
For their discovery of grid cells, May-Britt Moser, and Edvard Moser were awarded the
1535:
723:
344:
267:
259:
167:
1498:
1073:
1001:"Progressive increase in grid scale from dorsal to ventral medial entorhinal cortex"
853:
482:
109:
1173:
896:
533:
433:
254:
Neural activity in nearly every part of the hippocampal system is modulated by the
82:
1275:
37:
1377:
1343:"Accurate Path Integration in Continuous Attractor Network Models of Grid Cells"
296:
151:
1526:
1205:
606:"Direct recordings of grid-like neuronal activity in human spatial navigation"
311:
159:
155:
147:
130:
89:, and their students Torkel Hafting, Marianne Fyhn, and Sturla Molden at the
1514:
1436:
977:
931:
824:
Moser MB, Moser EI (1998). "Functional differentiation in the hippocampus".
1490:
1455:
1396:
1267:
1224:
1165:
1065:
1026:
985:
939:
888:
696:
639:
590:
525:
474:
425:
1324:
1122:
845:
810:
731:
17:
1057:
1041:
447:
Fyhn, M.; Hafting, T.; Witter, M. P.; Moser, E. I.; Moser, M. B. (2008).
1259:
1157:
678:
572:
517:
417:
121:
which can record single-neuron activity is implanted in the dorsomedial
1042:"Optimal Population Codes for Space: Grid Cells Outperform Place Cells"
1017:
1000:
880:
465:
448:
838:
10.1002/(SICI)1098-1063(1998)8:6<608::AID-HIPO3>3.0.CO;2-7
71:
50:
1482:
621:
1359:
301:
200:
108:
75:
36:
1341:
Burak, Y.; Fiete, I. R.; Sporns, O. (2009). Sporns, Olaf (ed.).
263:
67:
117:
To detect grid cell activity in a typical rat experiment, an
549:"A map of visual space in the primate entorhinal cortex"
547:
Killian, N. J.; Jutras, M. J.; Buffalo, E. A. (2012).
1040:
Mathis, A.; Herz, A. V. M.; Stemmler, M. B. (2012).
655:"Evidence for grid cells in a human memory network"
351:
List of distinct cell types in the adult human body
1410:Shrager, Y.; Kirwan, C. B.; Squire, L. R. (2008).
146:and Jonathon Dostrovsky reported the discovery of
951:
949:
1416:Proceedings of the National Academy of Sciences
653:Doeller, C. F.; Barry, C.; Burgess, N. (2010).
391:
389:
387:
385:
383:
381:
379:
377:
375:
373:
371:
369:
367:
365:
8:
1336:
1334:
93:(CBM) in Norway. They were awarded the 2014
1445:
1435:
1386:
1376:
1358:
1314:
1214:
1204:
1112:
1016:
800:
751:. Oxford University Press. Archived from
686:
629:
580:
464:
234:Interactions with hippocampal place cells
361:
321:The firing fields are organized into a
81:Grid cells were discovered in 2005 by
191:Nobel Prize in Physiology or Medicine
95:Nobel Prize in Physiology or Medicine
7:
170:as hypothesized by the psychologist
1307:10.1523/JNEUROSCI.17-15-05900.1997
1105:10.1523/JNEUROSCI.07-07-01951.1987
793:10.1523/JNEUROSCI.12-05-01945.1992
748:The Hippocampus as a Cognitive Map
164:The Hippocampus as a Cognitive Map
25:
1193:Frontiers in Systems Neuroscience
193:in 2014, alongside John O'Keefe.
30:For the concept in topology, see
1289:Samsonovich a, M. A. B. (1997).
91:Centre for the Biology of Memory
745:O'Keefe, J.; Nadel, L. (1978).
1087:Muller, RU; Kubie, JL (1987).
1:
310:In contrast to a hippocampal
1378:10.1371/journal.pcbi.1000291
1187:Kovács KA (September 2020).
724:10.1016/0006-8993(71)90358-1
1578:
1347:PLOS Computational Biology
29:
1206:10.3389/fnsys.2020.559186
256:hippocampal theta rhythm
1437:10.1073/pnas.0805414105
1295:Journal of Neuroscience
978:10.1126/science.1125572
932:10.1126/science.1099901
780:Journal of Neuroscience
138:Background of discovery
307:
206:
114:
42:
773:Quirk, M. R. (1992).
347:, discovered in 2008.
305:
204:
112:
40:
1527:Movie of a grid cell
1058:10.1162/NECO_a_00319
449:"Grid cells in mice"
306:A hexagonal lattice.
279:an agonist of GABA.
240:"complete remapping"
216:head direction cells
1471:Nature Neuroscience
1428:2008PNAS..10512034S
1369:2009PLSCB...5E0291B
1260:10.1038/nature06957
1252:2008Natur.453.1248H
1246:(7199): 1248–1252.
1158:10.1038/nature05601
1150:2007Natur.446..190F
970:2006Sci...312..758S
924:2004Sci...305.1258F
918:(5688): 1258–1264.
679:10.1038/nature08704
671:2010Natur.463..657D
610:Nature Neuroscience
573:10.1038/nature11587
565:2012Natur.491..761K
518:10.1038/nature10583
510:2011Natur.479..103Y
418:10.1038/nature03721
410:2005Natur.436..801H
210:dorsocaudal medial
1520:2019-04-15 at the
1046:Neural Computation
1018:10.1002/hipo.20504
881:10.1002/hipo.20114
466:10.1002/hipo.20472
308:
283:Possible functions
268:medial septal area
207:
115:
43:
32:Grid cell topology
1562:Spatial cognition
1301:(15): 5900–5920.
1144:(7132): 190–194.
1011:(12): 1200–1212.
964:(5774): 758–762.
665:(7281): 657–661.
504:(7371): 103–107.
459:(12): 1230–1238.
404:(7052): 801–806.
323:hexagonal lattice
276:attractor network
250:Theta rhythmicity
212:entorhinal cortex
123:entorhinal cortex
55:entorhinal cortex
16:(Redirected from
1569:
1503:
1502:
1466:
1460:
1459:
1449:
1439:
1407:
1401:
1400:
1390:
1380:
1362:
1338:
1329:
1328:
1318:
1286:
1280:
1279:
1235:
1229:
1228:
1218:
1208:
1184:
1178:
1177:
1133:
1127:
1126:
1116:
1084:
1078:
1077:
1052:(9): 2280–2317.
1037:
1031:
1030:
1020:
996:
990:
989:
953:
944:
943:
907:
901:
900:
864:
858:
857:
821:
815:
814:
804:
787:(5): 1945–1963.
770:
764:
763:
761:
760:
742:
736:
735:
707:
701:
700:
690:
650:
644:
643:
633:
601:
595:
594:
584:
544:
538:
537:
493:
487:
486:
468:
444:
438:
437:
393:
290:path integration
272:phase precession
244:"rate remapping"
172:Edward C. Tolman
127:action potential
21:
1577:
1576:
1572:
1571:
1570:
1568:
1567:
1566:
1532:
1531:
1522:Wayback Machine
1511:
1506:
1483:10.1038/nn.2901
1468:
1467:
1463:
1422:(33): 12034–8.
1409:
1408:
1404:
1353:(2): e1000291.
1340:
1339:
1332:
1288:
1287:
1283:
1237:
1236:
1232:
1186:
1185:
1181:
1135:
1134:
1130:
1086:
1085:
1081:
1039:
1038:
1034:
998:
997:
993:
955:
954:
947:
909:
908:
904:
866:
865:
861:
823:
822:
818:
772:
771:
767:
758:
756:
744:
743:
739:
709:
708:
704:
652:
651:
647:
622:10.1038/nn.3466
603:
602:
598:
559:(7426): 761–4.
546:
545:
541:
495:
494:
490:
446:
445:
441:
395:
394:
363:
359:
341:
285:
266:input from the
252:
236:
199:
140:
104:Euclidean space
87:May-Britt Moser
35:
28:
23:
22:
15:
12:
11:
5:
1575:
1573:
1565:
1564:
1559:
1554:
1549:
1544:
1534:
1533:
1530:
1529:
1524:
1510:
1509:External links
1507:
1505:
1504:
1477:(10): 1330–7.
1461:
1402:
1330:
1281:
1230:
1179:
1128:
1099:(7): 1951–68.
1079:
1032:
991:
945:
902:
875:(7): 841–852.
859:
816:
765:
737:
718:(1): 171–175.
712:Brain Research
702:
645:
616:(9): 1188–90.
596:
539:
488:
439:
360:
358:
355:
354:
353:
348:
340:
337:
333:
332:
329:
325:
319:
284:
281:
251:
248:
235:
232:
198:
195:
168:cognitive maps
139:
136:
97:together with
27:Type of neuron
26:
24:
14:
13:
10:
9:
6:
4:
3:
2:
1574:
1563:
1560:
1558:
1555:
1553:
1550:
1548:
1545:
1543:
1540:
1539:
1537:
1528:
1525:
1523:
1519:
1516:
1513:
1512:
1508:
1500:
1496:
1492:
1488:
1484:
1480:
1476:
1472:
1465:
1462:
1457:
1453:
1448:
1443:
1438:
1433:
1429:
1425:
1421:
1417:
1413:
1406:
1403:
1398:
1394:
1389:
1384:
1379:
1374:
1370:
1366:
1361:
1356:
1352:
1348:
1344:
1337:
1335:
1331:
1326:
1322:
1317:
1312:
1308:
1304:
1300:
1296:
1292:
1285:
1282:
1277:
1273:
1269:
1265:
1261:
1257:
1253:
1249:
1245:
1241:
1234:
1231:
1226:
1222:
1217:
1212:
1207:
1202:
1198:
1194:
1190:
1183:
1180:
1175:
1171:
1167:
1163:
1159:
1155:
1151:
1147:
1143:
1139:
1132:
1129:
1124:
1120:
1115:
1110:
1106:
1102:
1098:
1094:
1090:
1083:
1080:
1075:
1071:
1067:
1063:
1059:
1055:
1051:
1047:
1043:
1036:
1033:
1028:
1024:
1019:
1014:
1010:
1006:
1002:
995:
992:
987:
983:
979:
975:
971:
967:
963:
959:
952:
950:
946:
941:
937:
933:
929:
925:
921:
917:
913:
906:
903:
898:
894:
890:
886:
882:
878:
874:
870:
863:
860:
855:
851:
847:
843:
839:
835:
832:(6): 608–19.
831:
827:
820:
817:
812:
808:
803:
798:
794:
790:
786:
782:
781:
776:
769:
766:
755:on 2011-03-24
754:
750:
749:
741:
738:
733:
729:
725:
721:
717:
713:
706:
703:
698:
694:
689:
684:
680:
676:
672:
668:
664:
660:
656:
649:
646:
641:
637:
632:
627:
623:
619:
615:
611:
607:
600:
597:
592:
588:
583:
578:
574:
570:
566:
562:
558:
554:
550:
543:
540:
535:
531:
527:
523:
519:
515:
511:
507:
503:
499:
492:
489:
484:
480:
476:
472:
467:
462:
458:
454:
450:
443:
440:
435:
431:
427:
423:
419:
415:
411:
407:
403:
399:
392:
390:
388:
386:
384:
382:
380:
378:
376:
374:
372:
370:
368:
366:
362:
356:
352:
349:
346:
345:Boundary cell
343:
342:
338:
336:
330:
326:
324:
320:
317:
316:
315:
313:
304:
300:
298:
293:
291:
282:
280:
277:
273:
269:
265:
261:
257:
249:
247:
245:
241:
233:
231:
227:
225:
219:
217:
213:
203:
196:
194:
192:
187:
183:
181:
175:
173:
169:
165:
161:
157:
153:
149:
145:
137:
135:
132:
128:
124:
120:
111:
107:
105:
100:
96:
92:
88:
84:
79:
77:
73:
69:
65:
61:
56:
52:
49:is a type of
48:
39:
33:
19:
1515:Mosers Group
1474:
1470:
1464:
1419:
1415:
1405:
1350:
1346:
1298:
1294:
1284:
1243:
1239:
1233:
1196:
1192:
1182:
1141:
1137:
1131:
1096:
1092:
1082:
1049:
1045:
1035:
1008:
1004:
994:
961:
957:
915:
911:
905:
872:
868:
862:
829:
825:
819:
784:
778:
768:
757:. Retrieved
753:the original
747:
740:
715:
711:
705:
662:
658:
648:
613:
609:
599:
556:
552:
542:
501:
497:
491:
456:
452:
442:
401:
397:
334:
309:
294:
286:
253:
243:
239:
237:
228:
223:
220:
208:
188:
184:
179:
176:
163:
144:John O'Keefe
141:
116:
99:John O'Keefe
83:Edvard Moser
80:
46:
44:
1005:Hippocampus
869:Hippocampus
826:Hippocampus
453:Hippocampus
297:hippocampus
260:cholinergic
156:place field
152:hippocampus
150:in the rat
148:place cells
53:within the
1547:Perception
1536:Categories
1093:J Neurosci
759:2009-11-05
357:References
312:place cell
197:Properties
160:Lynn Nadel
131:place cell
18:Grid cells
1552:Neurology
1360:0811.1826
264:GABAergic
119:electrode
47:grid cell
1518:Archived
1499:17158393
1491:21909090
1456:18687893
1397:19229307
1268:18480753
1225:33013334
1166:17322902
1074:15755674
1066:22594833
1027:19021257
986:16675704
940:15333832
889:16145692
854:32384692
697:20090680
640:23912946
591:23103863
526:22051680
483:25196865
475:18683845
426:15965463
339:See also
142:In 1971
1542:Neurons
1447:2575247
1424:Bibcode
1388:2632741
1365:Bibcode
1325:9221787
1316:6573219
1248:Bibcode
1216:7511719
1174:4431307
1146:Bibcode
1123:3612226
1114:6568940
966:Bibcode
958:Science
920:Bibcode
912:Science
897:7237113
846:9882018
811:1578279
802:6575876
732:5124915
688:3173857
667:Bibcode
631:3767317
582:3565234
561:Bibcode
534:1978069
506:Bibcode
434:4405184
406:Bibcode
162:called
72:monkeys
1497:
1489:
1454:
1444:
1395:
1385:
1323:
1313:
1276:597353
1274:
1266:
1240:Nature
1223:
1213:
1199:: 68.
1172:
1164:
1138:Nature
1121:
1111:
1072:
1064:
1025:
984:
938:
895:
887:
852:
844:
809:
799:
730:
695:
685:
659:Nature
638:
628:
589:
579:
553:Nature
532:
524:
498:Nature
481:
473:
432:
424:
398:Nature
224:et al.
180:et al.
76:humans
74:, and
51:neuron
1557:Brain
1495:S2CID
1355:arXiv
1272:S2CID
1170:S2CID
1070:S2CID
893:S2CID
850:S2CID
530:S2CID
479:S2CID
430:S2CID
328:2007)
1487:PMID
1452:PMID
1393:PMID
1321:PMID
1264:PMID
1221:PMID
1162:PMID
1119:PMID
1062:PMID
1023:PMID
982:PMID
936:PMID
885:PMID
842:PMID
807:PMID
728:PMID
693:PMID
636:PMID
587:PMID
522:PMID
471:PMID
422:PMID
262:and
68:bats
64:mice
60:rats
1479:doi
1442:PMC
1432:doi
1420:105
1383:PMC
1373:doi
1311:PMC
1303:doi
1256:doi
1244:453
1211:PMC
1201:doi
1154:doi
1142:446
1109:PMC
1101:doi
1054:doi
1013:doi
974:doi
962:312
928:doi
916:305
877:doi
834:doi
797:PMC
789:doi
720:doi
683:PMC
675:doi
663:463
626:PMC
618:doi
577:PMC
569:doi
557:491
514:doi
502:479
461:doi
414:doi
402:436
1538::
1493:.
1485:.
1475:14
1473:.
1450:.
1440:.
1430:.
1418:.
1414:.
1391:.
1381:.
1371:.
1363:.
1349:.
1345:.
1333:^
1319:.
1309:.
1299:17
1297:.
1293:.
1270:.
1262:.
1254:.
1242:.
1219:.
1209:.
1197:14
1195:.
1191:.
1168:.
1160:.
1152:.
1140:.
1117:.
1107:.
1095:.
1091:.
1068:.
1060:.
1050:24
1048:.
1044:.
1021:.
1009:18
1007:.
1003:.
980:.
972:.
960:.
948:^
934:.
926:.
914:.
891:.
883:.
873:15
871:.
848:.
840:.
828:.
805:.
795:.
785:12
783:.
777:.
726:.
716:34
714:.
691:.
681:.
673:.
661:.
657:.
634:.
624:.
614:16
612:.
608:.
585:.
575:.
567:.
555:.
551:.
528:.
520:.
512:.
500:.
477:.
469:.
457:18
455:.
451:.
428:.
420:.
412:.
400:.
364:^
85:,
78:.
70:,
66:,
62:,
45:A
1501:.
1481::
1458:.
1434::
1426::
1399:.
1375::
1367::
1357::
1351:5
1327:.
1305::
1278:.
1258::
1250::
1227:.
1203::
1176:.
1156::
1148::
1125:.
1103::
1097:7
1076:.
1056::
1029:.
1015::
988:.
976::
968::
942:.
930::
922::
899:.
879::
856:.
836::
830:8
813:.
791::
762:.
734:.
722::
699:.
677::
669::
642:.
620::
593:.
571::
563::
536:.
516::
508::
485:.
463::
436:.
416::
408::
34:.
20:)
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.