826:. They also help in determining which nutrients are limiting in a localized system, if there is a limiting nutrient. The ratio can also be used to understand the formation of phytoplankton blooms and subsequently hypoxia by comparing the ratio between different regions, such as a comparison of the Redfield Ratio of the Mississippi River to the ratio of the northern Gulf of Mexico. Controlling N:P could be a means for sustainable reservoir management. It may even be the case that the Redfield Ratio is applicable to terrestrial plants, soils, and soil microbial biomass, which would inform about limiting resources in terrestrial ecosystems. In a study from 2007, soil and microbial biomass were found to have a consistent C:N:P ratios of 186:13:1 and 60:7:1, respectively on average at a global scale.
809:
residence times of these elements are greater than the mixing times of the oceans (~ 1000 years) can result in the ratio of nitrate to phosphate in the ocean interior remaining fairly uniform. It has been shown that phytoplankton play a key role in helping maintain this ratio. As organic matter sinks both nitrate and phosphate are released into the ocean via remineralization. Microorganisms preferentially consume oxygen in nitrate over phosphate leading to deeper oceanic waters having an N:P ratio of less than 16:1. From there, the ocean's currents upwell the nutrients to the surface where phytoplankton will consume the excess
Phosphorus and maintain a N:P ratio of 16:1 by consuming N
853:
stoichiometry. Also, when phosphorus is scarce, phytoplankton communities can lower their P content, raising the N:P. Additionally, the accumulation and quantity of dead phytoplankton and detritus can affect the availability of certain food sources which in turn affects the composition of the cell. In some ecosystems, the
Redfield ratio has also been shown to vary significantly by the dominant phytoplankton taxa present in an ecosystem, even in systems with abundant nutrients. Consequently, the system-specific Redfield ratio could serve as a proxy for plankton community structure.
62:, and empirically found the ratio to be C:N:P = 106:16:1. While deviations from the canonical 106:16:1 ratio have been found depending on phytoplankton species and the study area, the Redfield ratio has remained an important reference to oceanographers studying nutrient limitation. A 2014 paper summarizing a large data set of nutrient measurements across all major ocean regions spanning from 1970 to 2010 reported the global median C:N:P to be 163:22:1.
87:
664:
935:(cell walls). As a result of this, the Redfield-Brzezinski nutrient ratio was proposed for diatoms and stated to be C:Si:N:P = 106:15:16:1. Extending beyond primary production itself, the oxygen consumed by aerobic respiration of phytoplankton biomass has also been shown to follow a predictable proportion to other elements. The O
789:
one of the mechanisms initially proposed by
Redfield). However, subsequent modeling of feedback mechanisms, specifically nitrate-phosphorus coupling fluxes, do support his proposed mechanism of biotic feedback equilibrium, though these results are confounded by limitations in our current understanding of nutrient fluxes.
792:
In the ocean, a large portion of the biomass is found to be nitrogen-rich plankton. Many of these plankton are consumed by other plankton biomass which have similar chemical compositions. This results in a similar N:P ratio, on average, for all the plankton throughout the world’s oceans, empirically
754:
II) An equilibrium between seawater and planktonic nutrient pools is maintained through biotic feedback mechanisms. Redfield proposed a thermostat like scenario in which the activities of nitrogen fixers and denitrifiers keep the nitrate to phosphate ratio in the seawater near the requirements in the
788:
Laboratory experiments under controlled chemical conditions have found that phytoplankton biomass will conform to the
Redfield ratio even when environmental nutrient levels exceed them, suggesting that ecological adaptation to oceanic nutrient ratios is not the only governing mechanism (contrary to
843:
of phytoplankton grown under nitrogen or phosphorus limitation shows that this N:P ratio can vary anywhere from 6:1 to 60:1. While understanding this problem, Redfield never attempted to explain it with the exception of noting that the N:P ratio of inorganic nutrients in the ocean interior was an
808:
That the nitrate to phosphate ratio in the interior of all of the major ocean basins is highly similar is possibly due to the residence times of these elements in the ocean relative to the ocean's circulation time, roughly 100 000 years for phosphorus and 2000 years for nitrogen. The fact that the
907:
in the ocean. Since then experimentation has proven that Iron is a limiting factor for primary production. Iron-rich solution was added to 64 km area which led to an increase in phytoplankton primary production. As a result an extended
Redfield ratio was developed to include this as part of
847:
Although the
Redfield ratio is remarkably stable in the deep ocean, it has been widely shown that phytoplankton may have large variations in the C:N:P composition, and their life strategy plays a role in the C:N:P ratio. This variability has made some researchers speculate that the Redfield ratio
852:
ratio fundamentally present in both prokaryotes and eukaryotes, which contributes to it being the most common composition. There are several possible explanations for the observed variability in C:N:P ratios. The speed at which the cell grows has an influence on cell composition and thereby its
758:
In 1958, almost a quarter century after first discovering the ratios, Redfield leaned toward the latter mechanism in his manuscript, The
Biological Control of Chemical Factors in the Environment. Redfield proposed that the ratio of nitrogen to phosphorus in plankton resulted in the global ocean
821:
The research that resulted in this ratio has become a fundamental feature in the understanding of the biogeochemical cycles of the oceans, and one of the key tenets of biogeochemistry. The
Redfield ratio is instrumental in estimating carbon and nutrient fluxes in
17:
743:
Redfield’s analysis of the empirical data led to him to discover that across and within the three oceans and
Barents Sea, seawater had an N:P atomic ratio near 20:1 (later corrected to 16:1), and was very similar to the average N:P of phytoplankton.
775:
Redfield discovered the remarkable congruence between the chemistry of the deep ocean and the chemistry of living things such as phytoplankton in the surface ocean. Both have N:P ratios of about 16:1 in terms of atoms. When nutrients are not
1834:
Moore, C. M.; Mills, M. M.; Arrigo, K. R.; Berman-Frank, I.; Bopp, L.; Boyd, P. W.; Galbraith, E. D.; Geider, R. J.; Guieu, C.; Jaccard, S. L.; Jickells, T. D.; La Roche, J.; Lenton, T. M.; Mahowald, N. M.; Marañón, E. (September 2013).
912:
states that the ratio should be 106 C:16 N:1 P:0.1-0.001 Fe. The large variation for Fe is a result of the significant obstacle of ships and scientific equipment contaminating any samples collected at sea with excess Fe. It was this
750:
I) The N:P in plankton tends towards the N:P composition of seawater. Specifically, phytoplankton species with different N and P requirements compete within the same medium and come to reflect the nutrient composition of the seawater.
755:
protoplasm. Considering that at the time little was known about the composition of “protoplasm", or the bulk composition of phytoplankton, Redfield did not attempt to explain why its N:P ratio should be approximately 16:1.
834:
The
Redfield ratio was initially derived empirically from measurements of the elemental composition of plankton in addition to the nitrate and phosphate content of seawater collected from a few stations in the
784:
elemental ratio C:N:P in most phytoplankton is 106:16:1. Redfield thought it wasn't purely coincidental that the vast oceans would have a chemistry perfectly suited to the requirements of living organisms.
813:
via nitrogen fixation. While such arguments can potentially explain why the ratios are fairly constant, they do not address the question why the N:P ratio is nearly 16 and not some other number.
1386:
Levich, A. P. (1 February 1996). "The role of nitrogen-phosphorus ratio in selecting for dominance of phytoplankton by cyanobacteria or green algae and its application to reservoir management".
1699:
Arrigo, Kevin R.; Dunbar, Robert B.; Lizotte, Michael P.; Robinson, D. H. (2002). "Taxon-specific differences in C/P and N/P drawdown for phytoplankton in the Ross Sea, Antarctica".
848:
perhaps is a general average in the modern ocean rather than a fundamental feature of phytoplankton, though it has also been argued that it is related to a homeostatic protein-to-
860:
such as marine phytoplankton in an oceanic region do not conform to the canonical Redfield ratio, the fundamental concept of this ratio remains valid and useful.
58:
who in 1934 first described the relatively consistent ratio of nutrients in marine biomass samples collected across several voyages on board the research vessel
174:
20:
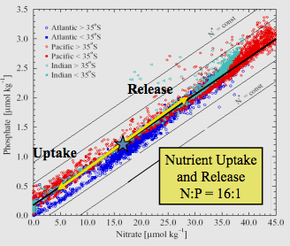
Relationship of phosphate to nitrate uptake for photosynthesis in various regions of the ocean. Note that nitrate is more often limiting than phosphate
459:
132:
1359:
Dodds, Walter (2006). "Nutrients and the "dead zone" : the link between nutrient ratios and dissolved oxygens in the Northern Gulf of Mexico".
1257:
632:
1530:
Loladze, Irakli; Elser, James J. (2011). "The origins of the Redfield nitrogen-to-phosphorus ratio are in a homoeostatic protein-to-rRNA ratio".
159:
1750:
Benner, R; Pakulski, JD; McCarthy, M; Hedges, JI; Hatcher, PG (1992). "Bulk chemical characteristics of dissolved organic matter in the ocean".
695:
513:
1644:
Martiny, Adam C.; Pham, Chau T. A.; Primeau, Francois W.; Vrugt, Jasper A.; Moore, J. Keith; Levin, Simon A.; Lomas, Michael W. (2013-03-17).
1572:
Klausmeier, C., Litchman, E., Daufresne, T. et al. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 429, 171–174 (2004).
839:. This was later supported by hundreds of independent measurements of dissolved nitrate and phosphate. However, the composition of individual
1917:
Brzezinski, Mark A. (1985). "THE Si:C:N RATIO OF MARINE DIATOMS: INTERSPECIFIC VARIABILITY AND THE EFFECT OF SOME ENVIRONMENTAL VARIABLES1".
637:
981:
793:
found to average approximately 16:1. When these organisms sink into the ocean interior, their biomass is consumed by bacteria that, in
102:
917:
that resulted in early evidence suggesting that iron concentrations were high and not a limiting factor in marine primary production.
1901:
740:, analyzing data for C, N, and P content in marine plankton, and referenced data collected by other researchers as early as 1898.
1965:
562:
1977:
567:
552:
232:
622:
520:
226:
220:
767:(16:1). He considered how the cycles of not just N and P but also C and O could interact to result in this match.
627:
247:
1372:
688:
948:
602:
530:
525:
508:
353:
214:
112:
896:
612:
127:
982:"On the proportions of organic derivatives in sea water and their relation to the composition of plankton"
432:
208:
169:
137:
681:
668:
617:
348:
1261:
1012:"Concentrations and ratios of particulate organic carbon, nitrogen, and phosphorus in the global ocean"
1848:
1759:
1708:
1657:
1598:
1488:
1309:
1221:
1169:
1120:
953:
909:
823:
544:
446:
202:
107:
781:
493:
309:
164:
1942:
1872:
1783:
1732:
1681:
1512:
1461:
1411:
1341:
1081:
904:
572:
340:
324:
319:
242:
55:
747:
To explain this phenomenon, Redfield initially proposed two mutually non-exclusive mechanisms:
1934:
1897:
1864:
1775:
1724:
1673:
1626:
1587:"A simple nutrient-dependence mechanism for predicting the stoichiometry of marine ecosystems"
1555:
1547:
1504:
1453:
1403:
1333:
1325:
1239:
1187:
1138:
1089:
1073:
1043:
503:
282:
117:
1064:
REDFIELD, ALFRED C. (1958). "The Biological Control of Chemical Factors in the Environment".
2010:
1926:
1856:
1814:
1767:
1716:
1665:
1616:
1606:
1539:
1496:
1445:
1395:
1368:
1317:
1229:
1177:
1128:
1033:
1023:
794:
469:
363:
358:
1646:"Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter"
86:
2015:
928:
900:
889:
597:
464:
397:
385:
314:
188:
1852:
1763:
1712:
1661:
1602:
1492:
1313:
1225:
1173:
1124:
1819:
1802:
1621:
1586:
1038:
1011:
836:
802:
717:
607:
557:
498:
412:
295:
290:
151:
2004:
1930:
1736:
1543:
1434:"C:N:P stoichiometry in soil: is there a "Redfield ratio" for the microbial biomass?"
914:
725:
52:
45:
1946:
1787:
1516:
1465:
1415:
1282:
1685:
1345:
924:
733:
721:
390:
304:
258:
196:
78:
1876:
1771:
1210:"Redfield revisited: 1. Regulation of nitrate, phosphate, and oxygen in the ocean"
16:
1891:
737:
729:
454:
427:
422:
417:
407:
377:
263:
122:
59:
1449:
1182:
1157:
402:
41:
1989:
1938:
1868:
1728:
1677:
1551:
1457:
1407:
1329:
1243:
1191:
1142:
1077:
1611:
1433:
1158:"On phytoplankton growth rates and particulate C: N: P ratios at low light1"
932:
869:
857:
764:
713:
1779:
1630:
1559:
1508:
1337:
1093:
1047:
1836:
1028:
1720:
1297:
1234:
1209:
1010:
Martiny, Adam C.; Vrugt, Jasper A.; Lomas, Michael W. (9 December 2014).
777:
37:
1971:. University of Hawaii School of Ocean and Earth Science and Technology.
1500:
1321:
1085:
736:, Redfield participated in several voyages on board the research vessel
1479:
Arrigo, KR (2005). "Marine microorganisms and global nutrient cycles".
1399:
920:
840:
798:
760:
709:
485:
268:
1645:
1573:
1133:
1108:
1860:
1669:
1298:"The polar ocean and glacial cycles in atmospheric CO2 concentration"
895:
In particular, iron (Fe) was considered of great importance as early
881:
873:
647:
33:
1803:"Marine bacteria and biogeochemical cycling of iron in the oceans"
642:
15:
1296:
Sigman, Daniel M.; Hain, Mathis P.; Haug, Gerald H. (July 2010).
1109:"Global patterns of marine nitrogen fixation and denitrification"
801:
the organic matter to form dissolved inorganic nutrients, mainly
885:
877:
849:
1373:
10.1890/1540-9295(2006)004[0211:NATDZT]2.0.CO;2
1801:
Tortell, PD; Maldonado, MT; Granger, J; Price, NM (1999).
1837:"Processes and patterns of oceanic nutrient limitation"
844:
average with small scale variability to be expected.
1432:Cleveland, Cory C.; Liptzin, Daniel (2007-09-01).
856:Despite reports that the elemental composition of
1427:
1425:
868:Some feel that there are other elements, such as
1591:Proceedings of the National Academy of Sciences
1059:
1057:
975:
973:
971:
969:
759:having a remarkably similar ratio of dissolved
1208:Lenton, Timothy M.; Watson, Andrew J. (2000).
1585:Galbraith, Eric D.; Martiny, Adam C. (2015).
1203:
1201:
1107:Gruber, Nicolas; Sarmiento, Jorge L. (1997).
708:For his 1934 paper, Alfred Redfield analyzed
689:
8:
1990:"MARINE BIOGEOCHEMISTRY: ON REDFIELD RATIOS"
830:Deviations from the canonical Redfield ratio
1983:. LSU School of the Coast and Environment.
1978:"Nutrient Stoichiometry - Redfield Ratios"
696:
682:
69:
1896:. Lamont-Doherty Geological Observatory.
1818:
1620:
1610:
1233:
1181:
1132:
1037:
1027:
1361:Frontiers in Ecology and the Environment
1890:Broecker, WS; Peng, T; Beng, Z (1982).
965:
939::C ratio has been measured at 138:106.
633:Territorialisation of carbon governance
77:
899:hypothesized that iron may also be a
638:Total Carbon Column Observing Network
7:
1258:"Green Ocean Project Nitrogen cycle"
1988:P.G. Falkowski; C.S. Davis (2004).
1574:https://doi.org/10.1038/nature02454
1388:Journal of Aquatic Ecosystem Health
1820:10.1111/j.1574-6941.1999.tb00593.x
32:is the consistent atomic ratio of
14:
1931:10.1111/j.0022-3646.1985.00347.x
1544:10.1111/j.1461-0248.2010.01577.x
888:which are also important in the
663:
662:
85:
48:and throughout the deep oceans.
989:James Johnstone Memorial Volume
51:The term is named for American
598:Climate reconstruction proxies
1:
1772:10.1126/science.255.5051.1561
923:need, among other nutrients,
1701:Geophysical Research Letters
1214:Global Biogeochemical Cycles
1113:Global Biogeochemical Cycles
568:Carbonate compensation depth
233:Particulate inorganic carbon
2032:
1162:Limnology and Oceanography
805:, nitrate, and phosphate.
623:Carbon capture and storage
227:Particulate organic carbon
221:Dissolved inorganic carbon
1450:10.1007/s10533-007-9132-0
1183:10.4319/lo.1986.31.6.1358
1156:Goldman, Joel C. (1986).
897:biological oceanographers
824:global circulation models
628:Carbon cycle re-balancing
1976:Lentz, Jennifer (2010).
949:Ecological stoichiometry
908:this balance. This new
603:Carbon-to-nitrogen ratio
563:Carbonate–silicate cycle
531:Carbon dioxide clathrate
526:Clathrate gun hypothesis
354:Net ecosystem production
215:Dissolved organic carbon
1612:10.1073/pnas.1423917112
1283:"Chemical Sensor Group"
864:Extended Redfield ratio
613:Deep Carbon Observatory
73:Part of a series on the
433:Continental shelf pump
209:Total inorganic carbon
175:Satellite measurements
30:Redfield stoichiometry
21:
1029:10.1038/sdata.2014.48
980:Redfield, AC (1934).
618:Global Carbon Project
349:Ecosystem respiration
19:
1966:"Biogeochemistry IV"
1919:Journal of Phycology
1807:Microbiology Ecology
1721:10.1029/2002GL015277
1235:10.1029/1999GB900065
1072:(3): 205–221, 230A.
954:Biogeochemical cycle
910:stoichiometric ratio
447:Carbon sequestration
203:Total organic carbon
1994:ScienceWeek. Nature
1853:2013NatGe...6..701M
1764:1992Sci...255.1561B
1713:2002GeoRL..29.1938A
1662:2013NatGe...6..279M
1603:2015PNAS..112.8199G
1501:10.1038/nature04159
1493:2005Natur.437..349A
1322:10.1038/nature09149
1314:2010Natur.466...47S
1226:2000GBioC..14..225L
1174:1986LimOc..31.1358G
1125:1997GBioC..11..235G
494:Atmospheric methane
460:Soil carbon storage
310:Reverse Krebs cycle
165:Ocean acidification
1964:Johnson, Zackary.
1893:Tracers in the Sea
1400:10.1007/BF00691729
1066:American Scientist
905:primary production
573:Great Calcite Belt
521:Aerobic production
341:Carbon respiration
283:Metabolic pathways
243:Primary production
56:Alfred C. Redfield
22:
1841:Nature Geoscience
1707:(19): 44–1–44-4.
1650:Nature Geoscience
1597:(27): 8199–8204.
1134:10.1029/97GB00077
706:
705:
504:Methane emissions
160:In the atmosphere
2023:
1997:
1984:
1982:
1972:
1970:
1951:
1950:
1914:
1908:
1907:
1887:
1881:
1880:
1861:10.1038/ngeo1765
1831:
1825:
1824:
1822:
1798:
1792:
1791:
1758:(5051): 1561–4.
1747:
1741:
1740:
1696:
1690:
1689:
1670:10.1038/ngeo1757
1641:
1635:
1634:
1624:
1614:
1582:
1576:
1570:
1564:
1563:
1527:
1521:
1520:
1487:(7057): 349–55.
1476:
1470:
1469:
1429:
1420:
1419:
1383:
1377:
1376:
1356:
1350:
1349:
1293:
1287:
1286:
1279:
1273:
1272:
1270:
1269:
1260:. Archived from
1254:
1248:
1247:
1237:
1205:
1196:
1195:
1185:
1168:(6): 1358–1363.
1153:
1147:
1146:
1136:
1104:
1098:
1097:
1061:
1052:
1051:
1041:
1031:
1007:
1001:
1000:
998:
996:
986:
977:
698:
691:
684:
671:
666:
665:
470:pelagic sediment
364:Soil respiration
359:Photorespiration
89:
70:
44:found in marine
2031:
2030:
2026:
2025:
2024:
2022:
2021:
2020:
2001:
2000:
1987:
1980:
1975:
1968:
1963:
1960:
1955:
1954:
1916:
1915:
1911:
1904:
1889:
1888:
1884:
1833:
1832:
1828:
1800:
1799:
1795:
1749:
1748:
1744:
1698:
1697:
1693:
1643:
1642:
1638:
1584:
1583:
1579:
1571:
1567:
1532:Ecology Letters
1529:
1528:
1524:
1478:
1477:
1473:
1438:Biogeochemistry
1431:
1430:
1423:
1385:
1384:
1380:
1358:
1357:
1353:
1308:(7302): 47–55.
1295:
1294:
1290:
1281:
1280:
1276:
1267:
1265:
1256:
1255:
1251:
1207:
1206:
1199:
1155:
1154:
1150:
1106:
1105:
1101:
1063:
1062:
1055:
1016:Scientific Data
1009:
1008:
1004:
994:
992:
984:
979:
978:
967:
962:
945:
938:
929:biogenic silica
901:limiting factor
890:ocean chemistry
866:
832:
819:
812:
773:
732:. As a Harvard
702:
661:
654:
653:
652:
592:
584:
583:
582:
547:
537:
536:
535:
488:
478:
477:
476:
465:Marine sediment
449:
439:
438:
437:
398:Solubility pump
386:Biological pump
380:
370:
369:
368:
343:
333:
332:
331:
315:Carbon fixation
300:
285:
275:
274:
273:
254:
238:
191:
189:Forms of carbon
181:
180:
179:
154:
144:
143:
142:
97:
68:
12:
11:
5:
2029:
2027:
2019:
2018:
2013:
2003:
2002:
1999:
1998:
1985:
1973:
1959:
1958:External links
1956:
1953:
1952:
1925:(3): 347–357.
1909:
1902:
1882:
1847:(9): 701–710.
1826:
1793:
1742:
1691:
1656:(4): 279–283.
1636:
1577:
1565:
1538:(3): 244–250.
1522:
1471:
1444:(3): 235–252.
1421:
1378:
1367:(4): 211–217.
1351:
1288:
1274:
1249:
1220:(1): 225–248.
1197:
1148:
1119:(2): 235–266.
1099:
1053:
1002:
964:
963:
961:
958:
957:
956:
951:
944:
941:
936:
865:
862:
837:Atlantic Ocean
831:
828:
818:
815:
810:
803:carbon dioxide
772:
769:
704:
703:
701:
700:
693:
686:
678:
675:
674:
673:
672:
656:
655:
651:
650:
645:
640:
635:
630:
625:
620:
615:
610:
608:Deep biosphere
605:
600:
594:
593:
590:
589:
586:
585:
581:
580:
578:Redfield ratio
575:
570:
565:
560:
558:Nutrient cycle
555:
549:
548:
545:Biogeochemical
543:
542:
539:
538:
534:
533:
528:
523:
518:
517:
516:
511:
501:
499:Methanogenesis
496:
490:
489:
484:
483:
480:
479:
475:
474:
473:
472:
462:
457:
451:
450:
445:
444:
441:
440:
436:
435:
430:
425:
420:
415:
413:Microbial loop
410:
405:
400:
395:
394:
393:
382:
381:
376:
375:
372:
371:
367:
366:
361:
356:
351:
345:
344:
339:
338:
335:
334:
330:
329:
328:
327:
322:
312:
307:
301:
299:
298:
296:Chemosynthesis
293:
291:Photosynthesis
287:
286:
281:
280:
277:
276:
272:
271:
266:
261:
255:
253:
252:
251:
250:
239:
237:
236:
230:
224:
218:
212:
206:
200:
193:
192:
187:
186:
183:
182:
178:
177:
172:
167:
162:
156:
155:
152:Carbon dioxide
150:
149:
146:
145:
141:
140:
135:
130:
125:
120:
115:
110:
105:
99:
98:
95:
94:
91:
90:
82:
81:
75:
74:
67:
64:
26:Redfield ratio
13:
10:
9:
6:
4:
3:
2:
2028:
2017:
2014:
2012:
2009:
2008:
2006:
1995:
1991:
1986:
1979:
1974:
1967:
1962:
1961:
1957:
1948:
1944:
1940:
1936:
1932:
1928:
1924:
1920:
1913:
1910:
1905:
1903:9780961751104
1899:
1895:
1894:
1886:
1883:
1878:
1874:
1870:
1866:
1862:
1858:
1854:
1850:
1846:
1842:
1838:
1830:
1827:
1821:
1816:
1812:
1808:
1804:
1797:
1794:
1789:
1785:
1781:
1777:
1773:
1769:
1765:
1761:
1757:
1753:
1746:
1743:
1738:
1734:
1730:
1726:
1722:
1718:
1714:
1710:
1706:
1702:
1695:
1692:
1687:
1683:
1679:
1675:
1671:
1667:
1663:
1659:
1655:
1651:
1647:
1640:
1637:
1632:
1628:
1623:
1618:
1613:
1608:
1604:
1600:
1596:
1592:
1588:
1581:
1578:
1575:
1569:
1566:
1561:
1557:
1553:
1549:
1545:
1541:
1537:
1533:
1526:
1523:
1518:
1514:
1510:
1506:
1502:
1498:
1494:
1490:
1486:
1482:
1475:
1472:
1467:
1463:
1459:
1455:
1451:
1447:
1443:
1439:
1435:
1428:
1426:
1422:
1417:
1413:
1409:
1405:
1401:
1397:
1393:
1389:
1382:
1379:
1374:
1370:
1366:
1362:
1355:
1352:
1347:
1343:
1339:
1335:
1331:
1327:
1323:
1319:
1315:
1311:
1307:
1303:
1299:
1292:
1289:
1285:. 2015-08-20.
1284:
1278:
1275:
1264:on 2015-04-02
1263:
1259:
1253:
1250:
1245:
1241:
1236:
1231:
1227:
1223:
1219:
1215:
1211:
1204:
1202:
1198:
1193:
1189:
1184:
1179:
1175:
1171:
1167:
1163:
1159:
1152:
1149:
1144:
1140:
1135:
1130:
1126:
1122:
1118:
1114:
1110:
1103:
1100:
1095:
1091:
1087:
1083:
1079:
1075:
1071:
1067:
1060:
1058:
1054:
1049:
1045:
1040:
1035:
1030:
1025:
1022:(1): 140048.
1021:
1017:
1013:
1006:
1003:
990:
983:
976:
974:
972:
970:
966:
959:
955:
952:
950:
947:
946:
942:
940:
934:
930:
926:
922:
918:
916:
915:contamination
911:
906:
902:
898:
893:
891:
887:
883:
879:
875:
871:
863:
861:
859:
854:
851:
845:
842:
838:
829:
827:
825:
816:
814:
806:
804:
800:
796:
790:
786:
783:
779:
770:
768:
766:
762:
756:
752:
748:
745:
741:
739:
735:
731:
727:
723:
719:
716:data for the
715:
711:
699:
694:
692:
687:
685:
680:
679:
677:
676:
670:
660:
659:
658:
657:
649:
646:
644:
641:
639:
636:
634:
631:
629:
626:
624:
621:
619:
616:
614:
611:
609:
606:
604:
601:
599:
596:
595:
588:
587:
579:
576:
574:
571:
569:
566:
564:
561:
559:
556:
554:
553:Marine cycles
551:
550:
546:
541:
540:
532:
529:
527:
524:
522:
519:
515:
512:
510:
507:
506:
505:
502:
500:
497:
495:
492:
491:
487:
482:
481:
471:
468:
467:
466:
463:
461:
458:
456:
453:
452:
448:
443:
442:
434:
431:
429:
426:
424:
421:
419:
416:
414:
411:
409:
406:
404:
401:
399:
396:
392:
389:
388:
387:
384:
383:
379:
374:
373:
365:
362:
360:
357:
355:
352:
350:
347:
346:
342:
337:
336:
326:
323:
321:
318:
317:
316:
313:
311:
308:
306:
303:
302:
297:
294:
292:
289:
288:
284:
279:
278:
270:
267:
265:
262:
260:
257:
256:
249:
246:
245:
244:
241:
240:
234:
231:
228:
225:
222:
219:
216:
213:
210:
207:
204:
201:
198:
195:
194:
190:
185:
184:
176:
173:
171:
168:
166:
163:
161:
158:
157:
153:
148:
147:
139:
136:
134:
133:Boreal forest
131:
129:
126:
124:
121:
119:
116:
114:
111:
109:
106:
104:
101:
100:
93:
92:
88:
84:
83:
80:
76:
72:
71:
65:
63:
61:
57:
54:
53:oceanographer
49:
47:
46:phytoplankton
43:
39:
35:
31:
27:
18:
1993:
1922:
1918:
1912:
1892:
1885:
1844:
1840:
1829:
1810:
1806:
1796:
1755:
1751:
1745:
1704:
1700:
1694:
1653:
1649:
1639:
1594:
1590:
1580:
1568:
1535:
1531:
1525:
1484:
1480:
1474:
1441:
1437:
1394:(1): 55–61.
1391:
1387:
1381:
1364:
1360:
1354:
1305:
1301:
1291:
1277:
1266:. Retrieved
1262:the original
1252:
1217:
1213:
1165:
1161:
1151:
1116:
1112:
1102:
1069:
1065:
1019:
1015:
1005:
993:. Retrieved
988:
925:silicic acid
919:
894:
867:
855:
846:
833:
820:
807:
797:conditions,
791:
787:
774:
757:
753:
749:
746:
742:
734:physiologist
707:
577:
391:Martin curve
378:Carbon pumps
305:Calvin cycle
259:Black carbon
197:Total carbon
138:Geochemistry
79:Carbon cycle
50:
29:
25:
23:
771:Explanation
730:Barents Sea
728:oceans and
455:Carbon sink
418:Viral shunt
408:Marine snow
264:Blue carbon
118:Deep carbon
113:Atmospheric
103:Terrestrial
2005:Categories
1268:2015-03-27
960:References
931:for their
927:to create
428:Whale pump
423:Jelly pump
403:Lipid pump
128:Permafrost
96:By regions
42:phosphorus
1939:1529-8817
1869:1752-0894
1737:129704137
1729:1944-8007
1678:1752-0894
1552:1461-0248
1458:1573-515X
1408:1573-5141
1330:0028-0836
1244:1944-9224
1192:1939-5590
1143:1944-9224
1078:0003-0996
933:frustules
870:potassium
858:organisms
765:phosphate
714:phosphate
66:Discovery
1947:86568341
1813:(1): 1.
1788:28604425
1780:17820170
1631:26056296
1560:21244593
1517:62781480
1509:16163345
1466:51898417
1416:84469052
1338:20596012
1094:24545739
1086:27827150
1048:25977799
995:March 1,
943:See also
778:limiting
738:Atlantis
718:Atlantic
669:Category
60:Atlantis
38:nitrogen
2011:Ecology
1849:Bibcode
1760:Bibcode
1752:Science
1709:Bibcode
1686:5677709
1658:Bibcode
1622:4500256
1599:Bibcode
1489:Bibcode
1346:4424883
1310:Bibcode
1222:Bibcode
1170:Bibcode
1121:Bibcode
1039:4421931
921:Diatoms
841:species
799:oxidize
795:aerobic
761:nitrate
726:Pacific
710:nitrate
514:Wetland
486:Methane
269:Kerogen
170:Removal
2016:Ratios
1945:
1937:
1900:
1877:249514
1875:
1867:
1786:
1778:
1735:
1727:
1684:
1676:
1629:
1619:
1558:
1550:
1515:
1507:
1481:Nature
1464:
1456:
1414:
1406:
1344:
1336:
1328:
1302:Nature
1242:
1190:
1141:
1092:
1084:
1076:
1046:
1036:
884:, and
882:copper
874:sulfur
780:, the
722:Indian
667:
648:CO2SYS
509:Arctic
248:marine
108:Marine
34:carbon
1981:(PDF)
1969:(PDF)
1943:S2CID
1873:S2CID
1784:S2CID
1733:S2CID
1682:S2CID
1513:S2CID
1462:S2CID
1412:S2CID
1342:S2CID
1082:JSTOR
991:: 176
985:(PDF)
782:molar
643:C4MIP
591:Other
235:(PIC)
229:(POC)
223:(DIC)
217:(DOC)
211:(TIC)
205:(TOC)
1935:ISSN
1898:ISBN
1865:ISSN
1776:PMID
1725:ISSN
1674:ISSN
1627:PMID
1556:PMID
1548:ISSN
1505:PMID
1454:ISSN
1404:ISSN
1334:PMID
1326:ISSN
1240:ISSN
1188:ISSN
1139:ISSN
1090:PMID
1074:ISSN
1044:PMID
997:2019
903:for
886:iron
878:zinc
850:rRNA
817:Uses
712:and
199:(TC)
123:Soil
40:and
24:The
1927:doi
1857:doi
1815:doi
1768:doi
1756:255
1717:doi
1666:doi
1617:PMC
1607:doi
1595:112
1540:doi
1497:doi
1485:437
1446:doi
1396:doi
1369:doi
1318:doi
1306:466
1230:doi
1178:doi
1129:doi
1034:PMC
1024:doi
763:to
28:or
2007::
1992:.
1941:.
1933:.
1923:21
1921:.
1871:.
1863:.
1855:.
1843:.
1839:.
1811:29
1809:.
1805:.
1782:.
1774:.
1766:.
1754:.
1731:.
1723:.
1715:.
1705:29
1703:.
1680:.
1672:.
1664:.
1652:.
1648:.
1625:.
1615:.
1605:.
1593:.
1589:.
1554:.
1546:.
1536:14
1534:.
1511:.
1503:.
1495:.
1483:.
1460:.
1452:.
1442:85
1440:.
1436:.
1424:^
1410:.
1402:.
1390:.
1363:.
1340:.
1332:.
1324:.
1316:.
1304:.
1300:.
1238:.
1228:.
1218:14
1216:.
1212:.
1200:^
1186:.
1176:.
1166:31
1164:.
1160:.
1137:.
1127:.
1117:11
1115:.
1111:.
1088:.
1080:.
1070:46
1068:.
1056:^
1042:.
1032:.
1018:.
1014:.
987:.
968:^
892:.
880:,
876:,
872:,
724:,
720:,
325:C4
320:C3
36:,
1996:.
1949:.
1929::
1906:.
1879:.
1859::
1851::
1845:6
1823:.
1817::
1790:.
1770::
1762::
1739:.
1719::
1711::
1688:.
1668::
1660::
1654:6
1633:.
1609::
1601::
1562:.
1542::
1519:.
1499::
1491::
1468:.
1448::
1418:.
1398::
1392:5
1375:.
1371::
1365:4
1348:.
1320::
1312::
1271:.
1246:.
1232::
1224::
1194:.
1180::
1172::
1145:.
1131::
1123::
1096:.
1050:.
1026::
1020:1
999:.
937:2
811:2
697:e
690:t
683:v
Text is available under the Creative Commons Attribution-ShareAlike License. Additional terms may apply.