This article may require cleanup to meet Knowledge's quality standards. The specific problem is: Inconsistent use of extinction dagger symbol. Please help improve this article if you can. (March 2024) (Learn how and when to remove this message) |
This is an incomplete list that briefly describes vertebrates that were extant during the Campanian, a stage of the Late Cretaceous Period which extended from 83.6 to 72.1 million years before present. Animals that lived in the Campanian include:
Cartilaginous fish
Chimaeriformes
| Chimaeras of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Campanian – Rupelian | Russia | Amylodon is an extinct genus of chimaera, it was considered to be a member of Chimaeridae but other studies have assigned it to either "Edaphodontidae" or Rhinochimaeridae. |  |
|
Cretaceous–Neogene | North America | Edaphodon is an extinct genus of chimaera. It is sometimes placed in the "Edaphodontidae", an unclearly defined group of chimaera with an uncertain position within the clade. | |
|
Cretaceous–Paleogene | North America | Elasmodus is an extinct genus of chimaera. It is sometimes placed in the "Edaphodontidae". | |
|
Middle Jurassic – Miocene. | Russia; USA | Ischyodus is an extinct genus of chimaera. It is sometimes placed in the "Edaphodontidae", while other authors place it into the extant Callorhinchidae. | |
†Hybodontiformes
| Hybodonts of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Ladinian–Maastrichtian |
|
Lonchidion selachos and Lonchidion griffisi were among the latest surviving species of hybodont. Lonchidion species were originally included to the closely related but earlier genus of Lissodus. |  |
|
Aptian–Maastrichtian | Montana and Wyoming, USA | Meristodonoides montanensis was one of the latest surviving species of hybodont, it was, along with the type species of its genus, originally described as a species of the earlier genus Hybodus. | |
|
Early-Late Cretaceous, 112–85 mya | North America | Ptychodus is an extinct genus of very large durophagous hybodont fish. | |
Selachimorpha
| Sharks of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Santonian to Campanian | Anglo-Paris Basin, France and the United Kingdom | Adnetoscyllium is an extinct monotypic genus of bamboo shark. It can be told apart from most other related genera via its lack of lateral cusplets. |    |
|
Early Cretaceous to Eocene | Sweden | Anomotodon is an extinct genus of shark related to the extant goblin shark. | |
|
Campanian | Judith River Formation, Montana; Bearpaw Formation, Alberta | Archaeotriakis is an extinct genus of false catshark. It is known from two species, mainly from isolated teeth. | |
|
Campanian-present | Northumberland Formation, British Columbia, Canada; Angola | A genus of chlamydoselachid sharks, it includes the modern frilled shark and Southern African frilled shark. | |
|
Late Cretaceous-present | Europe | Carcharias is a genus of mackerel sharks belonging to the family Odontaspididae. Once bearing many prehistoric species, all have gone extinct with the exception of the critically endangered sand tiger shark. | |
|
Cenomanian to present | North America; Europe | A genus of bamboo shark. Many species survive to this day. | |
|
Late Albian-Lutetian (103.13–46.25 mya) | North Africa; Middle East; North America | Cretalamna is a genus of extinct medium to large-sized shark in the family Otodontidae. It is considered by many to be the ancestor of the largest sharks to have ever lived. | |
|
Hauterivian-Maastrichtian | Aguja Formation, Mexico; Dinosaur Park Formation, Alberta; Judith River Formation, Saskatchewan, Canada and Montana, United States; Mesaverde Group, Wyoming; Åsen and Ignaberga, Sweden | Cretorectolobus is an extinct carpet shark. The type species is C. olsoni, which existed during the Campanian in Canada and the United States. | |
|
Albian-Campanian
107.59–73.2 mya |
North America; Sweden | Cretoxyrhina was one of the largest sharks of its time. It had convergently evolved similar appearance and build to the modern great white shark but it was only distantly related within Lamniformes. It was an apex predator in its ecosystem and preyed on a large variety of marine animals including mosasaurs, plesiosaurs, sharks and other large fish, pterosaurs, and occasionally dinosaurs. Cretoxyrhina was also among the fastest-swimming sharks, with hydrodynamic calculations suggesting burst speeds of up to 70 km/h. | |
|
Late Campanian | Angola; Northumberland Formation, British Columbia, Canada; Antarctica. | A very large chlamydoselachid shark and one of the biggest species of Hexanchiformes to have ever lived, might had surpassed lengths of 7 meters. | |
|
Campanian to Selandian | Northumberland Formation, Canada; Angola; New Zealand | Komoksodon is an extinct genus of hexanchiform shark. It is the only member of the monotypic family Komoksodontidae. | |
|
Early Jurassic–Eocene | Montana, North America; Sweden | Paraorthacodus is an extinct genus of shark. It a member of the family Paraorthacodontidae, though it was formerly regarded as a member of the family Palaeospinacidae, which is either placed in Hexanchiformes or in Synechodontiformes. It is known from over a dozen named species spanning from the Early Jurassic to the Paleocene, or possibly Eocene. Almost all members of the genus are exclusively known from isolated teeth, with the exception of P. jurensis from the Late Jurassic-Early Cretaceous of Europe, which is known from full body fossils from the Late Jurassic of Germany, which suggest that juveniles had a robust body with a round head, while adults had large body sizes with a fusiform profile. | |
|
Cenomanian-Maastrichtian | Middle East; Europe; North America | Pseudocorax is an extinct genus of mackerel sharks. It contains six valid species. It was formerly assigned to the family Anacoracidae, but is now placed in its own family Pseudocoracidae | |
|
Campanian-Late Miocene | Angola; Northumberland Formation, British Columbia, Canada; Antarctica. | A genus of chlamydoselachid sharks, many species, like the Campanian R. goliath and R. ludvigseni, reached length of above 4 meters. | |
|
Albian-Miocene | Egypt; Syria; North America | Scapanorhynchus is an extinct genus of mitsukurinid shark that lived from the early Cretaceous until possibly the Miocene if S. subulatus is a and not a sand shark. Their extreme similarities to the living goblin shark, Mitsukurina owstoni, lead some experts to consider reclassifying it as Scapanorhynchus owstoni. | |
|
Late Cretaceous–Paleocene | Europe; North America | Serratolamna is an extinct genus of mackerel sharks that is placed in the monotypic family Serratolamnidae. | |
|
Albian-Maastrichtian | Egypt; Europe; North America | Squalicorax, commonly known as the "crow shark", is a genus of extinct lamniform shark. Squalicorax was a medium-sized shark, typically measuring about 1.8–3 metres (5.9–9.8 ft) long. The largest specimen of S. pristodontus was signigicantly larger, measuring up to 4.8 metres (16 ft) long. | |
| Late Cretaceous | Aguja and Pen Formations of Big Bend National Park, Texas | Squatirhina is an extinct genus of cartilaginous fish, possibly an orectolobid carpet shark. | ||
|
Late Triassic–Palaeocene | USA; Europe | Synechodus is an extinct genus of shark belonging to the family Palaeospinacidae and order Synechodontiformes. It is known from 16 species primarily spanning from the Late Triassic to Paleocene. | |
Batoidea
| Rays of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Middle Santonian to early Campanian | Southern England | A species of ray in the family Ganopristidae of the suborder Sclerorhynchoidei. Sclerorhynchoids are an extinct suborder of rajiforms that superficially resembled the distantly related sawfishes due to convergence but were more closely related to skates. | 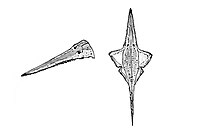 |
|
Lower Santonian to Campanian | Mesaverde Formation, Wyoming, USA; New Jersey, USA | A genus of sclerorhynchid sawskate. It is known only isolated rostral teeth. | |
|
Turonian-Thanetian
~89.3–55.8 mya |
Africa, the Middle East, North America, and South America | A genus of sclerorhynchoid ray. | |
|
Late Cretaceous, 94.3–66 mya | USA | An extinct genus of rajiform. | |
|
Late Cretaceous, 94.3–66 mya | USA | A genus of ptychotrygonid sclerorhynchoid. | |
|
Tithonian-present | A genus of rhinopristiform rays in the Rhinobatidae family. | ||
|
Santonian-Maastrichtian | Africa, the Middle East, North America, and South America | A genus of schizorhizid sclerorhynchoid. | |
|
Turonian-Maastrichtian | Africa, the Middle East, North America, and South America | A genus of ganopristid sclerorhynchoid. | |
Actinopterygii
| Basal ray-finned fish of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Campanian | Niobrara Formation, North America | Its type species, A. arcanus, is only known from poorly preserved single specimen, which makes hard to classify existing orders of Cretaceous actinopterygians. It was tentatively categorized as a palaeoniscoids but given its recent age and fragmentary nature this placement is unlikely. | ||
Chondrostei
| Chondrostei of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Late Cretaceous to present | North America | Acipenser is a genus of sturgeons. It is the largest genus in the order Acipenseriformes, with 17 living species, others are only known from fossil remains. At least one species, A. albertensis is known from the late Campanian. The genus is paraphyletic, containing all sturgeons that do not belong to Huso, Scaphirhynchus, or Pseudoscaphirhynchus, with many species more closely related to the other three genera than they are to other species of Acipenser. |
 | |
| Campanian (77.5 mya) | Judith River Formation, Montana, USA | Priscosturion, initially named Psammorhynchus, is an extinct genus of sturgeon known from one species, P. longipinnis. The fish belongs to the subfamily Priscosturioninae within the larger family Acipenseridae. | ||
Holostei
| Holostei of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Cretaceous – present | North America | A genus of amiids which contains all extant bowfin species. |
   | |
| Late Cretaceous – Eocene | North America | An extinct genus of amiids closely related to bowfin species. | ||
| Campanian to present | Americas | A genus of large lepisosteid gars with three extant species. | ||
| Campanian to present | North America, Brazil, France | A genus of lepisosteid gars. Multiple of its species are still extant. | ||
| Kirtlandian-Lancian (late Campanian-Maastrichtian) 74.8–66 Ma | Kirtland Formation, USA | A genus of large vidalamiin relative of bowfins. | ||
| Albian to Campanian | Tanout, Zinder, Niger | One of the latest surviving genera of Semionotiformes. | ||
Teleosteomorpha
| Teleosts and closely related fish of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Albian to Maastrichtian | France; United States | Anomoeodus is a genus of large pycnodontiform ray-finned fish belonging to the family Pycnodontidae. |
        | |
| Late Jurassic–Paleocene | Italy; North America | A genus of aspidorhynchiform fishes within the family Aspidorhynchidae. | ||
| Coniacian to Maastrichtian, 89–66 mya | Smoky Hill Member of the Niobrara Chalk, Pierre Shale, Mooreville Chalk, Demopolis Chalk, Wenonah Formation; North America | A genus of late surviving filter feeding pachycormid fish. It grew to at least 5 meters in total body length, substantially less than the related Leedsichthys from the Jurassic which likely grew up to 16.5 meters. | ||
| Cenomanian (99.6 ± 0.9 Mya and 93.5 ± 0.8 Mya) to the Maastrichtian (70.6 ± 0.6 Mya to 66 Mya) | North America; Europe | Cimolichthys is an extinct genus of 1.5- to 2.0-meter-long nektonic predatory aulopiform fish. Although the closest living relatives of Cimolichthys are lancetfish and lizardfish, the living animals would have resembled very large freshwater pikes. | ||
| Campanian to Maastrichtian, possibly to Paleocene | Mesa Verde Formation, USA | Coriops is an extinct genus of prehistoric bony fish. Might be an eel, an elopiform, a bonefish or an osteoglossomorph. | ||
| Campanian | Dactylopogon is an extinct genus of prehistoric elopiform fish. | |||
| Campanian | Westphalia, Germany | Echidnocephalus troscheli is an extinct, prehistoric halosaur. Fossil specimens suggest an animal already looking very much like modern halosaurs. | ||
| Late Cretaceous-Eocene | Black Creek Group, North Carolina | An extinct genus of bonefish fish in the family Phyllodontidae. | ||
| Cenomanian–Campanian | UK; Levant | A genus of extinct bony fish, possibly a halosaur. | ||
| Albian-Maastrichtian, ~112.6–66 Ma
|
North America; Egypt; Israel | An extinct genus of aulopiform ray-finned fish related to lancetfish and lizardfish. | ||
| Campanian-Maastrichtian | Angola and Nigeria | A genus of extinct durophagous elopiform fish in the suborder Albuloidea. | ||
| Santonian to Campanian | Alberta | An extinct genus of freshwater pikes. | ||
| Campanian to early Danian | Chaunaca Formation, Bolivia | A genus of prehistoric clupeomorph fish that is distantly related to modern anchovies and herrings. It contains one species, G. branisai. It inhabited freshwater or estuarine habitats. Its taxonomic identity was long uncertain, often being placed as a clupeid or an indeterminate clupeomorph, but more recent studies have placed it with the Ellimmichthyiformes. | ||
| 100–66 mya | North America | A genus of ichthyodectiform fish in the family Ichthyodectidae. Unlike its larger relative, Xiphactinus, by which it was preyed upon, it was a filter feeder instead of a large prey predator. | ||
| Campanian | Hornby Island, British Columbia, Canada | Gwawinapterus beardi is a species of saurodontid ichthyodectiform fish initially described as a very late-surviving member of the pterosaur family Istiodactylidae. | ||
| Coniacian to Campanian | North America | A genus of large ichthyodectiform fish in the family Ichthyodectidae. Like its larger close relative Xiphactinus was a voracious predator. | ||
| Campanian | Mooreville Chalk Formation, Alabama | An extinct genus of crossognathiform fish in the family Plethodidae. | ||
| Campanian to Maastrichtian | Alberta | An extinct genus of freshwater pikes. | ||
| Cenomanian–Maastrichtian | USA | An extinct genus of crossognathiform fish in the family Pachyrhizodontidae. | ||
| Late Cretaceous-Early Eocene, 99.7–40.4 mya | North America | An extinct genus of bonefish fish in the family Phyllodontidae. | ||
| Campanian | North America | An extinct genus of prehistoric elopiform fish in the family Elopidae. | ||
| Cenomanian–Campanian | North America | An extinct genus of crossognathiform fish in the family Plethodidae. Pentanogmius evolutus is among the largest known tselfatiiform fish with a length of over 1.7 meters. | ||
| Campanian | Sendenhorst, Germany | Sardinioides is an extinct genus of prehistoric myctophiform fish, and considered a sister group to Paleogene and modern neoscopelids and myctophids. | ||
| Campanian | Germany | Sardinius is an extinct genus of prehistoric ray finned fish, potentially a myctophiform. | ||
| Cenomanian–Campanian | North America | An extinct genus of prehistoric elopiform fish. | ||
| Cenomanian–Campanian | Sendenhorst, Germany | An extinct genus of aulopiform fish in Cheirothricidae, a family of fish capable of gliding flight over water, like the superficially similar flying fish, but more closely related to lizardfish. | ||
| Albian to Maastrichtian | North America; Europe; Australia; Venezuela | A genus of very large ichthyodectiform fish in the family Ichthyodectidae that could reach up to 4 meters in length. Xiphactinus was a voracious predator. | ||
Actinistia
| Coelacanths of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Lower Campanian possibly to early Maastrichtian | France | A species of averagely sized mawsoniid coelacanth. Related species of the same genus lived mostly on earlier stages of the Cretaceous. |
 |
|
Lower Campanian possibly to early Maastrichtian | USA | A very large latimerid coelacanth. It has been estimated to have been 3.5 – 4.5 meters in length. | |
Dipnoi
| Lungfish of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Olenekian-Eocene | Cosmpopolitan | A widespread genus of ceratodontiform lungfish in the family Ceratodontidae with multiple species throughout the mesozoic. |
  | |
| Norian-Maastrichtian | Malargüe Group, Argentina | A widespread genus of ceratodontiform lungfish in the family Ceratodontidae with multiple species throughout the mesozoic. | ||
| Albian to Recent | Brazil; Niger | A ceratodontiform lungfish in the family Neoceratodontidae. A species of this genus, the Australian Lungfish, survives to this day. | ||
| Campanian to Recent | Egypt; Sudan | A ceratodontiform lungfish in the family Protopteridae. Many species of this genus, survive to this day. | ||
| Albian–Maastrichtian | Egypt; Sudan | A genus of large ceratodontiform lungfish in the family Ceratodontidae. | ||
Amphibians
| †Amphibians of the Maastrichtian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
105.3–65.043 Ma, Albian to Maastrichtian | Canada
USA |
A salamander-like albanerpetontid that thrived in North America and Europe from the Early Cretaceous to the late Pliocene. |  |
|
Aptian-Campanian | Barun Goyot and Djadokhta Formation, Mongolia | Gobiates is an extinct genus of prehistoric frog. | |
|
Campanian | Dinosaur Park Formation, Alberta | A large sirenid, about the size of a hellbender. The palate is specialized for crushing, suggesting it may have fed on hard-bodied prey. | |
|
Campanian-Lancian | Dinosaur Park Formation, Alberta | An extinct genus of prehistoric salamanders in the family Batrachosauroididae. | |
|
Campanian-Lancian | Ellisdale Fossil Site, Marshalltown Formation, Monmouth County, New Jersey | Parrisia neocesariensis is an extinct species of batrachosauroidid salamander. | |
|
125–60.5 Ma, Aptian to Selandian | Hell Creek Formation, Montana, USA
Mexico
|
A little-known frog. | |
Lepidosaurs
| Lepidosaurs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
An agamid iguanian lizard. |
              | ||
|
A varanoid lizard of disputed affinity. | |||
|
A genus of fossil snakes in the extinct family of Madtsoiidae. | |||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanian lizard in the extinct clade Gobiguania. Compared to other iguanians, Anchaurosaurus has a relatively elongated skull, large eye sockets, and higher tooth crowns. | |||
| Brewster County, Aguja Formation, Texas, USA | A scincomorph. | |||
| 81 mya | Seonso Conglomerate Formation, South Korea | A genus of anguimorph lizard distantly related to the living Gila monster. | ||
| 84.9–70.6 mya | Bayan Mandahu Formation, Mongolia | A diploglossan anguimorph lizzard. | ||
| Brewster County, Aguja Formation, Texas, USA | A large, possibly durophagous, night lizard. | |||
| A basal anguimorph lizard closely related to the extant Xenosauridae. | ||||
| A varanoid lizard that resembles Aiolosaurus. | ||||
| A relatively small basal mosasaurine marine lizard. | ||||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanid iguanian lizard. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A scincomorph lizard. | |||
|
A genus of extinct varanoid lizards. | |||
| A genus of plioplatecarpine mosasaurs. | ||||
|
A genus of halisaurine mosasaurs. | |||
| Gobi Desert, Mongolia | A genus of anguimorph lizards, originally thought to be closely related to monitor lizards. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A priscagamid iguanian lizard. | |||
| Mongolia | A genus belonging to Polyglyphanodontia, the only major clade of lizards that went extinct during the K-T extinction event. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A scincomorph lizard. | |||
| Djadochta Formation, Gobi Desert, Mongolia | An anguimorph lizard that resembled a Gila monster though it was not necessarily closely related to it. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A teiid lacertoid lizard. | |||
|
A genus of medium-sized mosasaurine mosasaurs. Along with its closest relatives in the tribe Globidensini it had distinctive globular teeth. | |||
| An anguimorph lizard that resembled the modern Heloderma lizards. | ||||
| A comparatively small mosasaurs. Type genus of the subfamily Halisaurine. | ||||
| A madtsoiid snake. | ||||
| Djadochta Formation, Gobi Desert, Mongolia | A scincomorph lizard. | |||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanian lizard in the extinct clade Gobiguania. | |||
|
A sphenodontid rhynchocephalian closely related to the extant tuatara. | |||
|
Cerro Tortuga site near Lamarque, Allen Formation, Argentina | A basal neosphenodont rhynchocephalian. | ||
|
New Zealand, Japan, Antarctica | A large plioplatecarpine mosasaur. | ||
|
A madtsoiid snake. | |||
| A priscagamid iguanian lizard. Mimeosaurus is unique among iguanians in having premaxilla bones at the tip of the snout that are reduced in size, as well as having two pairs of enlarged canine-like teeth in the maxilla | ||||
|
A genus of large mosasaurine mosasaurs, contained some of the largest marine reptiles to have ever lived. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A basal lizard of contested affinities. Might be related to the contemporary Bainguis. | |||
|
An anguid anguimorph. | |||
|
†Ovoo
|
An early varanid. | |||
|
Late Cretaceous | North America | Palaeosaniwa is a platynotan anguimorph of uncertain affinities. It was among the largest terrestrial lizards known from the Mesozoic era, roughly comparable to a large monitor lizard in size, only exceeded by Asprosaurus and Chianghsia. | |
|
A varanoid lizard. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A scincomorph lizard. | |||
|
A madtsoiid snake. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A priscagamid iguanian lizard. | |||
| USA, possibly Belgium and Africa | A genus of plioplatecarpine mosasaurs. | |||
| Bayan Mandahu Formation Gobi Desert, Mongolia | Originally thought to be a priscagamid, it's sub-pleurodont dentition instead of acrodont permanent dentition indicates that it might had been a more basal iguanian, a potential "missing link" between Chamaeleontiformes and Pleurodonta. | |||
|
A genus of plioplatecarpine mosasaurs. | |||
| Djadochta Formation, Gobi Desert, Mongolia | A priscagamid iguanian lizard. | |||
|
A genus of large mosasaurine mosasaurs. | |||
|
An anguimorph lizard closely related to Monstersauria Varanoidea. | |||
| A madtsoiid snake. | ||||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanian lizard in the extinct clade Gobiguania. | |||
| A varanoid lizard. | ||||
| Djadochta Formation, Gobi Desert, Mongolia | It was originally proposed and argued that Sineoamphisbaenia was the oldest known amphisbaenian; this, however, was challenged and it's now considered to belong of the extinct clade Polyglyphanodontia. | |||
| Djadochta Formation, Gobi Desert, Mongolia | Previously thought to be a scincomorph, it was later placed in a clade containing Lacertoidea+Scincomorpha. A more recent description of the genus, placed it as a stem-group relative of amphisbaenians within Lacertoidea. | |||
| New Zealand, Japan, Antarctica | A medium-sized tylosaurine mosasaur. | |||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanian lizard in the extinct clade Gobiguania. | |||
| A varanoid lizard. | ||||
|
A genus of large tylosaurine mosasaurs, contained some of the largest marine reptiles to have ever lived. | |||
| Djadochta Formation, Gobi Desert, Mongolia | An iguanian lizard in the extinct clade Gobiguania. | |||
Testudines
| Turtles of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Cretaceous – Oligocene,
125–28 Mya |
North America | A large, freshwater pantryonychian turtle. |   | |
| Early Upper Cretaceous–Oligocene
94.3–28.4 Mya |
Germany | Allopleuron is a genus of extinct basal sea turtle, which measured 2-to-2.5-metre long in life. The type species is Allopleuron hofmanni. It is a basal member of the clade Pancheloniidae, closely related to Protosphargis. Similar to Protosphargis, it was characterized by shell reduction. | ||
| Campanian,
80.21–74.21 Mya |
Pierre Shale, South Dakota, USA | Archelon is a giant marine turtle in the extinct family Protostegidae. It is the largest turtle ever to have been sufficiently documented, with the biggest specimen measuring 4.6 m from head to tail and 2.2–3.2 t in body mass. | ||
| 83 Mya | New Jersey | An extinct large marine turtle in the family Protostegidae. Though known from single humerus its full size has been extrapolated as being 3 m. | ||
| Campanian-Maastrichtian,
~84–66 |
Horseshoe Canyon Formation, Oldman Formation and Dinosaur Park Formation, Alberta; Aguja Formation, Texas to Mexico | A large, terrestrial pantryonychian turtle in the family Nanhsiungchelyidae . It is considered to be the largest terrestrial turtle of its time. | ||
| Campanian–Maastrichtian | Selma Group, Alabama, USA | Calcarichelys is a small protostegid, with a carapace length of 25.3 cm. It is probably closely related to Chelosphargis, as they share some characteristics. Unlike Chelosphargis, Calcarichelys is characterized by thorn-like neural plates. | ||
| Campanian | Mooreville Chalk Formation (the lower part of the Selma Group), Alabama | Corsochelys halinches is an extinct species of basal dermochelyid sea turtle related to the modern leatherback turtle. | ||
| Late Cretaceous,
89–70 Ma |
Mooreville Chalk, central Alabama, United States | An extinct genus of stem-cheloniid sea-turtle. | ||
| Campanian | Cal Torrades locality, lower Perles Formation, Spanish Pyrenees | Leviathanochelys is an extinct genus of giant basal sea turtle. Although Leviathanochelys, at an extrapolated body length of 3.74 m, could reach similar proportions as the largest protostegids, it was more closely related to modern cheloniid sea turtles and possibly a sister taxon to the stem-cheloniid Allopleuron hoffmanni. | ||
| Early Campanian – Maastrichtian | Osoushinai Formation, Yezo Group, Hokkaido, Japan | Corsochelys halinches is an extinct species of basal dermochelyid sea turtle related to the modern leatherback turtle. | ||
| Late Cretaceous,
84.9–70.6 Mya |
Bearpaw Shale Formation, Alberta | Nichollsemys baieri is an extinct species of basal chelonioid sea turtles. | ||
| Campanian
83.5 Mya |
Smoky Hill Chalk formation, western Kansas; Mooreville Chalk Formation; Rybushka Formation, Saratov Oblast, Russia. | Protostega gigas is a species of giant marine turtle in the extinct family Protostegidae. It is one of the largest turtles to have ever lived. | ||
| Late Cretaceous,
~85–75 Ma |
Wangshi Group, Shandong, China, | A lindholmemydid pantestudinoid turtle. | ||
| Campanian
80 Mya |
Aguja Formation, Texas | An extinct marine turtle in the family Protostegidae. | ||
| Late Cretaceous | Smoky Hill Chalk, western Kansas | Toxochelys is an extinct genus of marine turtle. Toxochelys had carapace about 90 centimeters. Phylogenetic analysis shows that Toxochelys belong to an extinct lineage of turtles transitional between modern sea turtles and other turtles. | ||
†Plesiosaurs
| Plesiosaurs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| 73.5 Ma | Bearpaw Formation, Alberta. | Albertonectes is a genus of very large elasmosaurid plesiosaur with an extremely long neck. |      | |
| Coniacian–Campanian | North America | Dolichorhynchops is an extinct genus of polycotylid plesiosaur. | ||
| 80.5 mya | Pierre Shale, Kansas, USA | Elasmosaurus is a genus of elasmosaurid plesiosaur with a very long neck. | ||
| 77 mya | New Zealand | Mauisaurus is a dubious genus of elasmosaurid plesiosaur known from fragmentary remains. | ||
| Campanian, 80.5–75 mya | Sweden; Russia | Scanisaurus is a dubious genus of plesiosaur, most likely an elasmosaurid. It was relatively small and short necked compared to most elasmosaurids, sometimes been described as a "mesodiran" plesiosaur. | ||
| 83.5–80.5 mya | Logan County, Kansas | Styxosaurus is a genus of large elasmosaurid plesiosaur. | ||
| 73.5 Ma | Bearpaw Formation, Saskatchewan, Canada. | Terminonatator is a genus of elasmosaurid plesiosaur. | ||
†Choristoderans
| Choristoderans of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Campanian to Paleocene | North America | Champsosaurus is a genus of crocodile-like neochoristodere. |  | |
Crocodylomorphs
| Crocodylomorphs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
Early Campanian – Late Maastrichtian 83.5–66 Ma | Italy | Initially placed within Alligatoridae but has since been reclassified as a more basal globidontan, the oldest and most primitive known to date. Recent studies have since resolved Acynodon as a basal eusuchian crocodylomorph, outside of the Crocodylia crown group, and a close relative to Hylaeochampsa. |
     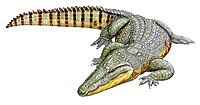
|
|
Early Campanian 83.5–80.5 mya | Kristianstad Basin, Sweden | A member of the extinct family Dyrosauridae, Aigialosaurus was a long- and narrow-snouted crocodylomorph. In contrast to modern crocodylians, which typically have long and slender teeth, the teeth of Aigialosuchus were stout and short. | |
|
76.9–75.8 Ma | Dinosaur Park Formation, Alberta | A basal member of Globidonta, a group of alligatoroids more derived than Deinosuchus or Leidyosuchus that includes living alligators and caimans. | |
|
Late Campanian-Maastrichtian | Spain | A genus of average-sized basal eusuchian. Might be synonymous with its contemporary relative genus Allodaposuchus. | |
|
84.9–66.043 Ma | Spain and France, | A genus of average-sized basal eusuchian, growing to around 3 m (9.8 ft) long. The main feature that distinguishes this species from other related crocodylomorphs is the orientation of a groove at the back of the skull called the cranioquadrate passage; unlike the cranioquadrate passages of other crocodylomorphs, which are only visible at the back of the skull, the cranioquadrate passage of this variety is visible when the skull is viewed from the side. The genus includes many species but it may be paraphyletic in regards to its contemporary relative genera Agaresuchus and Lohuecosuchus. | |
|
Campanian | Djadochta Formation, Mongolia | Its classification beyond that of a basal crocodylomorph is indeterminant because of the fragmentary nature of the material associated with the genus. | |
|
early Campanian to Danian | Hell Creek Formation, Montana, USA | A globidontan distinguished by an enlarged fourth maxillary tooth in the upper jaw. | |
|
82–73 Ma | USA | A basal alligatoroid and one of the largest crocodylomorphs that ever lived. | |
|
84.9–70.6 Ma | USA | Denazinosuchus is a genus of goniopholidid mesoeucrocodylian. | |
|
Early Campanian 75–71 Ma | Djadokhta Formation, Gobi Desert, Mongolia. | A member of Gobiosuchidae, a family of late surviving but basal non-mesoeucrocodilian crocodyliforms. | |
|
Middle Campanian | Alberta | A basal medium-sized alligatoroid. | |
|
Late Campanian-early Maastrichtian | Spain | A genus of average-sized basal eusuchian. Might be synonymous with its contemporary relative genus Allodaposuchus. | |
|
Late Cretaceous, ~96–70 Ma | Nemegt Formation, Mongolia | A paralligatorid mesoeucrocodylian. | |
|
83.6–72.1 mya | Djadochta Formation, Mongolia | A paralligatorid mesoeucrocodylian. | |
|
Zos Canyon, Gobi Desert, Mongolia. | A gobiosuchid. | ||
†Pterosaurs
| Pterosaurs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Campanian-Maastrichtian | Allen Formation, Patagonia, Argentina | A genus of moderately large azhdarchid pterosaurs. |
   | |
| Rybushka Formation, Petrovsk, Russia | A genus of moderately of medium to large sized azhdarchid pterosaurs. | |||
| USA, North America | Geosternbergia was originally a species of Pteranodon and is famous for its oddly shaped crest. | |||
| Montana, USA | Small azhdarchoid pterosaur, probably a tapejarid | |||
| New Mexico, USA, and Alberta, Canada | Known primarily from forearm elements; tentatively assigned to Azhdarchidae, though most likely not part of it. | |||
| mid-western United States | Nyctosaurus is a genus of pteranodontian pterosaur in the family Nyctosauridae. Nyctosaurids are characterized by their lack of all but the wing finger which may suggest that it spent almost all of its time in the air, rarely walking on the ground. | |||
| Montana, USA | Piksi is a genus of pterosaurs containing the single species Piksi barbarulna. | |||
| Kansas, USA, North America | Pteranodon is a genus of pterosaurs which included some of the largest known flying reptiles, with wingspans over 6 metres | |||
| Saratov, Russia | A large pterosaur, either an azhdarchoid, a pteranodontid or a nyctosaurid. | |||
Dinosaurs
During the Campanian age, a radiation among dinosaur species occurred. In North America, for example, the number of known dinosaur genera rises from 4 at the base of the Campanian to 48 in the upper part. This development is sometimes referred to as the "Campanian Explosion". However, it is not yet clear if the event is artificial, i.e. the low number of genera in the lower Campanian can be caused by a lower preservation chance for fossils in deposits of that age. The generally warm climates and large continental area covered in shallow sea during the Campanian probably favoured the dinosaurs. In the following Maastrichtian stage, the number of North American dinosaur genera found is 30% less than in the upper Campanian.
†Ornithischia
†Ankylosaurs
| Ankylosaurs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Point Loma Formation, California, USA | A medium-sized ankylosaurid, estimated to be around 6 m (20 ft) long. |  
| ||
| Santa Marta Formation, James Ross Island, Antarctica | A stocky ankylosaur protected by armor plates embedded in the skin. Although a complete skeleton has not been found, the species is estimated to have reached a maximum length of 4 meters (13 feet). Displays characteristics of both ankylosaurids and nodosaurids. | |||
| Campanian to Maastrichtian | Dinosaur Park Formation, Alberta, Canada | A bulky nodosaurid at roughly 6.6 m (22 ft) long. It had small, ridged bony plates on its back and many sharp spikes along its body sides. The four largest spikes jutted out from the shoulders on each side, two of which were split into subspines in some specimens. Its skull had a pear-like shape when viewed from above. | ||
| A dubious ankylosaurid, possibly a synonym of Pinacosaurus. | ||||
| 78.5 mya | Menefee Formation, New Mexico, USA | A nodosaurid. | ||
| Judith River Formation | known from a single tooth | |||
| Judith River Formation, Alberta, Canada; Montana, USA | A 5.5–7 m long nodosaurid. | |||
| 75 mya | Judith River Formation, Montana, USA | An ankylosaurine. | ||
†Theskelosaurids
| Parksosaurids of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
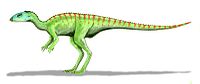
| 77–76 mya | Oldman Formation Alberta, Canada | A close relative of the contemporary Orodromeus. |  | |
| 81 mya | Seonso Conglomerate, Boseong, Korea | Koreanosaurus was likely a burrowing dinosaur. Unlike its orodromine relatives, Koreanosaurus is assumed to have been a quadruped. It reached 2–2.4 meters in body length. | ||
| 76.7 mya | Two Medicine Formation, Montana, USA | Orodromeus was a small fast bipedal herbivore that probably coexisted with dinosaurs such as Daspletosaurus and Einiosaurus. Its length was estimated by Horner & Weishampel at 2.5 metres. | ||
†Ceratopsians
| Ceratopsians of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
74.2 million years ago | Glacier County, Two Medicine Formation, Montana, U.S.A. | A highly derived centrosaurine ceratopsid in the tribe Pachyrhinosaurini. |         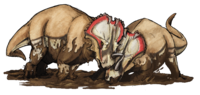  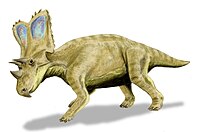                      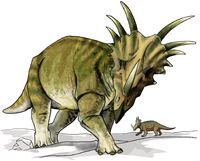        |
|
77 million years ago | Aguja Formation, Texas | A chasmosaurine ceratopsid. | |
|
middle Campanian-age, 77.5 mya | Oldman Formation, Alberta, Canada | Its size has been estimated at 5.8 metres and 3,500 kg, it was a rather large centrosaurine. | |
|
72–71 mya | Horseshoe Canyon Formation, Alberta | Anchiceratops ornatus was a medium-sized chasmosaurine with a distinctive frill. Fragmentary horn and frill remains from older Campanian formations in Alberta may indicate the existence of a second earlier species. | |
|
Late Campanian | Judith River Formation, Montana, USA | A basal centrosaurine ceratopsid. The holotype was small for a ceratopsid but it might had been a juvenile. | |
|
~72–71 mya | Barun Goyot Formation and Bayan Mandahu Formation, Mongolia | A rather small protoceratopsid. | |
|
75–71 mya | Djadochta Formation, Mongolia | A small dubious ceratopsian with affinities to Udanoceratops or potentially a juvenile Protoceratops. | |
|
74.5 | Two Medicine Formation, Montana, USA | A dubious centrosaurine known only from partial juvenile specimens. | |
|
Late Campanian | Barun Goyot Formation, Mongolia | A protoceratopsid known by a partial juvenile postcranial skeleton with skull. | |
|
76.5–75.5 mya | Dinosaur Park Formation, Alberta | A derived centrosaurine. | |
|
Two Medicine Formation, Montana, USA | A leptoceratopsid. | ||
|
||||
|
||||
|
||||
|
||||
|
A basal centrosaurine. | |||
|
79–74.9 mya | All 4 named species are known from the Judith River Formation of Montana, USA | A dubious genus of ceratopsid ceratopsians. | |
|
||||
|
||||
|
~75.9–75.3 | Judith River Formation, Montana, USA | A basal centrosaurine known from a nearly complete disarticulated skeleton of a moderately-sized subadult. | |
|
||||
|
77 mya | Oldman Formation, Alberta | ||
|
||||
|
||||
|
||||
|
||||
|
80.8 mya | Aguja Formation, Texas | A basal centrosaurine. | |
|
||||
|
Middle Campanian, 77.5 Ma | Judith River Formation, Montana, USA | A centrosaurine originally thought to be a chasmosaurine. | |
|
83.5–80 mya | Menefee Formation, New Mexico, USA | The oldest recognized centrosaurine ceratopsid. | |
|
||||
|
||||
|
||||
|
||||
|
75.0–73.4 mya | Kirtland Formation, New Mexico, USA | Navajoceratops was found to represent a stratigraphic and morphological intermediate between Pentaceratops and Anchiceratops. Navajoceratops was also found to be marginally less derived than Terminocavus. | |
|
Late Campanian, about 76.0–75.5 mya | Upper Kaiparowits Formation, Grand Staircase–Escalante National Monument, Utah, United States. | A basal centrosaurine. It had a pair of cattle-like horns above its eyes, the largest ones among Centrosaurinae. | |
|
Late Cretaceous, ~77–66 mya | Lago Colhué Huapí Formation, Chubut Province, Argentina | A dubious genus of possible ceratopsian affinity | |
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
75.0–74.6 mya | Kirtland Formation, New Mexico, USA | It was proposed that Terminocavus is a part of a long anagenetic lineage of chasmosaurs. The genera (in sequence) Utahceratops, Pentaceratops, Navajoceratops, Terminocavus, and Anchiceratops would, under this model, represent a single population of organisms changing in form over time, rather than as a diverse assemblage of close relatives. | |
|
75 mya | Southwestern USA | A very large chasmosaurine, either a synonym and an exceptionally large individual of Pentaceratops sternbergii or a basal triceratopsin. | |
|
||||
|
||||
|
||||
|
||||
|
79.5 mya | Foremost Formation, Alberta | A basal centrosaurine. | |
|
72 mya | Coahuila, Aguja Formation, Mexico | A centrosaurine. | |
†Pachycephalosaurs
| Pachycephalosaurs of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Middle Campanian,
80–77.5 mya |
Foremost Formation, Alberta | Colepiocephale lambei is a domed pachycephalosaur characterized by the lack of a lateral and posteriosquamosal shelf, a steeply down-turned parietal, and the presence of two incipient nodes tucked under the posterior margin of the parietosquamosal border. |
         | |
| Late Campanian,
75–73 Mya |
Dinosaur Park Formation, Belly River Group, Alberta | Foraminacephale was a small to medium-sized pachycephalosaur. | ||
| 76 mya | Boro Khovil, Djadochta Formation, Mongolia | Goyocephale is a primitive pachycephalosaurian, and was originally included in the family Homalocephalidae, which united the genus with Homalocephale, which also has a flat skull roof, however, many more recent phylogenetic analyses tend to find Homalocephalidae to be a paraphyletic family, with the genera included simply being consecutive branches sister to Pachycephalosauridae, or as consecutive branches within Pachycephalosauridae. Goyocephale is distinguished from Homalocephale by its overall proportions, the shape of its supratemporal fenestra, and heterodont dentition, although the two share multiple features. | ||
| Late Campanian,
75 mya |
Belly River Group, Alberta | There is some debate amongst paleontologists as to whether the animal represents a distinct genus, or if it is synonymous with Stegoceras. Some recent publications indicate it may be a valid genus. | ||
| Campanian, 77.5–75 mya | Belly River Group, Dinosaur Park Formation and Oldman Formation, Alberta; Judith River Formation, Montana | Distinguished from other pachycephalosaurs by having a depressed parietal region, wide frontoparietal dome, broad nasal characteristics, reduced prefontal lobes, and a reduced parietosquamosal shelf. A possible synonym of Stegoceras. | ||
| Late Cretaceous | Omnogov, Nemegt Formation, Mongolia | Sporting a flat, wedge-shaped skull roof, Homalocephale was different from other pachycephalosaurs. | ||
| Campanian | Dinosaur Park Formation, Alberta | Likely a synonym of Stegoceras. | ||
| Late Cretaceous | Djadochta Formation, Mongolia | It was similar in many ways to its close relative, Homalocephale but unlike the flattened wedge-shaped skull of later (a possible juvenile trait also potentially seen in early growth stages of Pachycephalosaurus), the head of Prenocephale was rounded and sloping. | ||
|
Campanian to Maastrichtian |
S. goodwini: Den-na-zin Member of the Kirtland Formation S. edmontonensis: Horseshoe Canyon Formation, Alberta S. lyonsi: Dinosaur Park Formation, Alberta
|
A highly derived genus of pachycephalosaurs with multiple species. | |
|
~77.5–74 mya |
S. validum: Dinosaur Park Formation and Oldman Formation, Alberta ?S. novomexicanum: Fruitland Formation and Kirtland Formation, New Mexico |
A genus of medium to small sized basal pachycephalosaurid. | |
| Late Campanian,
75–73 Mya |
Barun Goyot Formation, Mongolia | Tylocephale was a medium-sized pachycephalosaur. | ||
| 77 mya | Aguja Formation, Texas | A medium-sized pachycephalosaur. The cranial dome of Texacephale possessed five to six vertical flanges on each lateral side, connecting it with the postorbital bone. | ||
| Latest Campanian - Maastrichtian | Upper member of the Xiaoyan Formation, at the town of Yansi, Shexian County, Anhui Province | A genus of small basal pachycephalosaurian. Despite being one of the smallest non-avian dinosaurs found the fused bones in its skull suggest that it was an adult at death. | ||
†Ornithopods
| Ornithopods of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
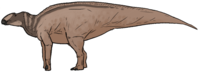
| 79 mya | Montana and Utah, USA | A genus of basal saurolophine hadrosaur. |
                            | |
| 78 mya | Wahweap Formation, Utah, US | A lambeosaurine, possibly closely related to Parasaurolophus. | ||
| 74 mya | Kirtland Formation, Texas | A saurolophine hadrosaur. | ||
| 76.9 mya | Aguja Formation, Texas | A lambeosaurine. | ||
| Aguja Formation, Texas | Formerly referred to Kritosaurus and Gryposaurus it might be a much more basal hadrosaurid. | |||
| Upper Santonian-Lower Campanian boundary 85.8 mya | Bostobe Formation, Kazakhstan | A basal lambeosaurine hadrosaur, Aralosaurus was about the size of an elephant. Although very little is known about Aralosaurus (only one near complete skull has been found); it was identified by a beak with nearly 1,000 small teeth in 30 rows. The back of an Aralosaurus skull was wide, a feature suggestive of large jaw muscles used to power its chewing apparatus. | ||
| 76.5 mya | Montana, USA; Alberta, Canada | Brachylophosaurus was a basal saurolophine hadrosaur which reached an adult length of 9 meters (30 feet). | ||
| 77–76.5 mya | Alberta, Canada | A derived lambeosaurine. Corythosaurus weighed in at 4 tonnes and measured roughly 10 metres (33 feet) from nose to tail. | ||
| 75 mya | Montana, USA | A dubious genus of hadrosaur known only from dental fossils. | ||
| 73.0–76.5 mya | Canada | Edmontosaurus, a saurolophine hadrosaur, included some of the largest ornithischian species, measuring up to 12 metres (39 feet) long and weighing around 4.0 metric tons (4.4 short tons). | ||
| 85 mya | Argentina | Gasparinisaura was a small ornithopod of the basal clade Elasmaria. In 2010 Gregory S. Paul estimated the length at 1.7 metres, the weight at thirteen kilogrammes. | ||
| 72 mya | Mongolia | A relatively large hadrosauromorph. | ||
| 83–75.5 mya | Alberta, Canada | Gryposaurus was a saurolophine hadrosaur of typical size and shape. | ||
| 79.5 mya | New Jersey, USA | One of the most basal hadrosaurids and the namesake genus of the family. It was likely bipedal for the purposes of running, but could use its forelegs to support itself while feeding. | ||
| 75-67 mya | Alberta, Canada | A derived lambeosaurine, Hypacrosaurus is most easily distinguished from other hollow-crested duckbills by its tall neural spines and the form of its crest. The neural spines, which project from the top of the vertebrae, are 5 to 7 times the height of the body of their respective vertebrae in the back, which would have given it a tall back in profile. The skull's hollow crest is like that of Corythosaurus, but is more pointed along its top, not as tall, wider side to side, and has a small bony point at the rear | ||
| North Carolina and Missouri, USA | A large hadrosauroid known only by a couple of vertebrae. | |||
| 73 mya | North America | The type specimen of Kritosaurus navajovius is only represented by a partial skull and lower jaws, and associated postcranial remains. | ||
| 76-75 mya | Alberta, Canada | A derived lambeosaurine hadrosaur, namesake genus of the subfamily. | ||
| 80 mya | Alabama, USA | A genus of hadrosauromorphs. | ||
| Aguja Formation, Texas | A basal hadrosaur. | |||
| 76.7 mya | Montana, USA | Maiasaura was a large basal saurolophine hadrosaur, attaining an adult length of about 9 metres (30 feet) and had the typical hadrosaurid flat beak and a thick nose. It had a small, spiky crest in front of its eyes. The crest may have been used in headbutting contests between males during the breeding season. | ||
| Late Campanian, 74–72 mya | Grès à Reptiles Formation, France. | A rhabdodontid. | ||
| China | A possibly dubious genus of hadrosaur known by a partial lower jaw from a juvenile. | |||
| Austria | A rhabdodontid. | |||
| 73 mya | New Mexico, USA | Naashoibitosaurus, a saurolophine hadrosaur, based as it is on a single partial skeleton, is not well known in terms of anatomy. Its skull, the most thoroughly described portion, has a low nasal crest that peaks in front of the eyes, but does not strongly arch as in Gryposaurus. | ||
| 80 mya | Sakhalin island | A lambeosaurine hadrosaur known from subadult remains. | ||
| 78 mya | New Mexico, USA | A genus of basal saurolophine hadrosaur. | ||
| 76.5–73 mya | Alberta, Canada; New Mexico and Utah, USA | A large derived lambeosaurine hadrosaur. | ||
| 79 mya | Montana, USA; Alberta, Canada | A large basal saurolophine hadrosaur which reached an adult length of over 9 meters (30 feet). | ||
| 76-75 mya | Alberta, Canada | Prosaurolophus was a large-headed duckbill; the most complete described specimen has a skull around 0.9 meters (3.0 feet) long on a ~8.5-meter long skeleton (~28 ft). It had a small, stout, triangular crest in front of the eyes; the sides of this crest were concave, forming depressions. The upper arm was relatively short. | ||
| 75 mya | Montana, USA | A dubious genus of lambeosaurine hadrosaurs. Historically, several species were assigned to it, all based on extremely fragmentary remains, but there is no evidence to support these assignments. | ||
| 72 mya | France; Spain; Haţeg Island, Romania | It is unclear whether it was an iguanodont or a hypsilophodont, and may be a "missing link" between the two. Current evidence indicates it is an iguanodont similar to Tenontosaurus. | ||
| 69.5–68.5 mya | North America, Asia | Saurolophus is known from material including nearly complete skeletons, giving researchers a clear picture of its bony anatomy. S. osborni, the rarer Albertan species, was around 9.8 meters (32 feet) long, with its skull a meter long (3.3 feet). Its weight is estimated at 1.9 tonnes (2.1 tons). S. angustirostris, the Mongolian species, was larger; the type skeleton is roughly 12 meters (39 feet) long, and larger remains are reported. | ||
| ~76–72 mya | Lago Colhué Huapí Formation, Golfo San Jorge Basin, Patagonia, Argentina | A genus of ornithopod dinosaur, possibly an elasmarian. Sektensaurus is the first known non-hadrosauroid ornithopod of central Patagonia. | ||
| 72 mya | China | A very large saurolophine hadrosaur closely related to Edmontosaurus, it is one of the longest and largest known hadrosaurids and possibly the largest non-sauropod dinosaur; the composite skeleton of a medium-sized individual mounted at the Geological Institute of China in Beijing measures 14.72 metres (48.3 feet) in length. | ||
| 76–75 mya | Dinosaur Park Formation, Alberta | A dubious genus of hadrosaurid dinosaur with a complicated taxonomic history. | ||
| 72–71 mya | China | A genus of hadrosauromorph dinosaur. | ||
| 77 mya | Montana, USA | Trachodon is a dubious genus of hadrosaurid dinosaur based on teeth. Numerous species had been referred to this genus later reassigned to different genera or proven dubious. | ||
| 72 mya | Southern China | A basal but relatively large lambeosaurine hadrosaur. | ||
| 72 mya | Mexico | A large derived lambeosaurine. | ||
†Sauropoda
| Sauropods of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| South America | Lago Colhué Huapi Formation, Argentina | A robust yet moderately sized derived titanosaur. Aelosaurus had a widespread distribution as its remains were found in multiple sites from South America. |
     | |
| Neuquén Province, Argentina | Andesaurus is a genus of basal titanosaurian sauropod dinosaur. | |||
| Lago Colhué Huapi Formation, Argentina | A moderately large basal titanosaur. Argyrosaurus is the only titanosaur known to possess carpals. | |||
| Cerro Fortaleza Formation, Argentina | Dreadnoughtus is one of the largest titanosaurs known. | |||
| Adamantina Formation and Cambabe Formation, Brazil | A fairly small titanosaur, only 7 meters long, a close relative of Aeolosaurus. | |||
| North East, China | A member of the Euhelopodidae sauropods. | |||
| Allen Formation and Anacleto Formation, both in Argentina; Palacio Formation, Uruguay | A rather large yet elegantly built titanosaur, potentially related to earlier lognkosaurs. | |||
| Santonian-Campanian | Bajo de la Carpa Formation, Argentina | A dubious genus of small titanosaur, likely a saltasaurid, known from only a single dorsal vertebra. | ||
| Anacleto Formation, Argentina | Small-sized saltasaurid titanosaur very closely related to the Asian genus Saltasaurus.
| |||
| Campanian-Maastrichtian | Allen Formation, Rio Negro Province, Argentina | As saltasaurid titanosaur closely related to the Asian genus Saltasaurus. At a length of only 8 meters it was one of the smallest known sauropods. | ||
Theropoda
David J. Varrichio observes that during the late Campanian Alberta and Montana had very similar theropods despite significant differences in the types of herbivorous dinosaur faunas.
| Non-avian theropods of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Allen Formation?, Anacleto Formation?, Argentina | An abelisaurid ceratosaur that probably reached 7 to 9 meters in length; known from only one partial skull. |
  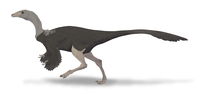                | ||
| A derived tyrannosauroid. | ||||
| An ornithomimid ornithomimosaur. | ||||
| A small dromaeosaurid. | ||||
| A large derived tyrannosauroid. | ||||
| A troodontid | ||||
| Dinosaur Park Formation, Alberta | A caenagnathid oviraptorosaur. | |||
| An oviraptorid oviraptorosaur. | ||||
| Dinosaur Park Formation, Alberta | An caenagnathid oviraptorosaur. It was substantially smaller than its contemporaneous relatives Caenagnathus and Chirostenotes. | |||
| A derived abelisaurid ceratosaur that probably reached 7 to 9 meters in length | ||||
| Dinosaur Park Formation, Alberta | A caenagnathid oviraptorosaur. | |||
|
A genus of large and robustly built tyrannosaurines. | |||
| Judith River Formation | A dubious tyrannosaurid known from tooth remains. | |||
| A small to medium-sized dromaeosaurid maniraptoran theropod with a sickle-like claw. | ||||
| An ornithomimid ornithomimosaur. | ||||
| A derived tyrannosaur. | ||||
| 78 mya | Menefee Formation, New Mexico, USA | A large basal tyrannosaurine. | ||
| A troodontid | ||||
| An albertosaurine tyrannosaurid. | ||||
| A basal ornithomimosaur. | ||||
| An oviraptorid oviraptorosaur. | ||||
|
†Kol |
An alvarezsaurid. | |||
| A large robust theropod of uncertain affinities, possibly a tyrannosaur. | ||||
| A caenagnathid oviraptorosaur. | ||||
| A dromaeosaurid. | ||||
| Wahweap Formation, Utah | A 7-meter tyrannosaurine known from a partially complete skull, some vertebrae and a complete pubis | |||
| A dromaeosaurid. | ||||
| A small halszkaraptorine maniraptoran. | ||||
| A relatively large-bodied therizinosaurid. | ||||
| A small derived ceratosaur. | ||||
| An ornithomimid ornithomimosaur. | ||||
| Mongolia | Oviraptor is an oviraptorid oviraptorosaur in the subfamily Oviraptorinae. | |||
| An alvarezsaur. | ||||
| Var, France | A small dromaeosaurid. | |||
| A coelurosaur form taxon known only from teeth and lower jaw fragments. | ||||
| A troodontid. | ||||
| A dromaeosaurid. | ||||
| An alvarezsaur. | ||||
| An ornithomimid ornithomimosaur. | ||||
| A basal tyrannosaurine. | ||||
| A close relative of Daspletosaurus. | ||||
| A troodontid. | ||||
| A dromaeosaurid. | ||||
| Var, France | A dromaeosaurid. | |||
| Mongolia and China | A dromaeosaurid. | |||
| Wangshi Group, Zhucheng, China | One of the largest tyrannosaurids at between 10 and 12 meters. Known from a lower jaw and maxilla slightly smaller than those of the later Tyrannosaurus. | |||
Avialae
| Birds of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| A genus of enantiornithine birds of uncertain affinities. |
    | |||
| A genus of basal ornithuran birds. | ||||
| A genus of basal ornithuromorph birds. | ||||
| A genus of flightless hesperornithiform birds. | ||||
| A genus of freshwater hesperornithiform birds. Although Hesperornithes probably lost their flight abilities by the end of the Early Cretaceous, minimal pachyostosis in Brodavis suggests the possibility of some volant abilities. | ||||
| A genus of flightless hesperornithid birds. | ||||
| An enantiornithe of uncertain affinities. It was estimated to have a wingspan of around 127–185 cm and may have been 75 to 110 cm long when fully grown, making it one of the largest known enantiornitheans to date. | ||||
| ~85 mya | Bostobe Formation, Kazakhstan | An extinct genus of birds. The type species, C. sarysuensis, is known only from an isolated, asymmetrical contour feather | ||
| Elsornis is a genus of basal enantiornithine birds. It was flightless or nearly so. | ||||
| A genus of large, primitive and potentially flightless avialan. | ||||
| An avisaurid enantiornithe, previously thought to be a species of the later closely related Maastrichtian genus Avisaurus. | ||||
| A genus of euenantiornithine enantiornithe. It possessed a robust beak which convergently evolved with those of modern birds. | ||||
| A genus of avisaurid enantiornithe. It lived in a coastal environment. | ||||
| A genus of flightless hesperornithid birds. | ||||
| A genus of small ground birds. It was either an enantiornithe or a basal ornithuromorph. | ||||
| A genus of toothy seabird-like ornithuran. | ||||
| A genus of avisaurid enantiornithe. Although closely related to some of the largest avisaurids, members of this genus were very small birds. | ||||
| Latest Campanian | Northumberland Formation, British Columbia, Canada. | A genus of large marine diving vegaviid bird with an estimated body mass of 1.5 kg. | ||
| A genus of avisaurid enantiornithes, closely related to the later genus Avisaurus. | ||||
| Limenavis is the closest known relative of modern birds. | ||||
| A marine bird from Chile. It had the midfeet of a foot-propelled diving bird, but its relationships are enigmatic. The only known species is from the Campanian-Maastrichtian boundary. | ||||
| A genus of basal long-tailed avialan. May potentially be a non-avialan genus of theropod. | ||||
Mammals
| Mammals of the Campanian | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Late Cretaceous | North America | A marsupialiform metatherian. | ||
| A marsupialiform metatherian. | ||||
| Djadokhta Formation, Mongolia | A multituberculate allotherian. | |||
| Bayan Mandahu Formation and Djadokhta Formation, Mongolia | An insectivorous leptictid eutherian. | |||
| Also known as Gobibaatar or Tugrigbaatar. It's a multituberculate allotherian. | ||||
| Type genus of Zalambdalestidae, a basal clade of Asian eutherians. | ||||
See also
- List of fossil sites (with link directory)
- List of vertebrate fauna of the Maastrichtian stage
References
- See Weishampel et al. (2004)
- ^ Brown, C. M.; Evans, D. C.; Ryan, M. J.; Russell, A. P. (2013). "New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta". Journal of Vertebrate Paleontology. 33 (3): 495. doi:10.1080/02724634.2013.746229. S2CID 129160518.
- ^ Min Huh; Dae-Gil Lee; Jung-Kyun Kim; Jong-Deock Lim; Pascal Godefroit (2011). "A new basal ornithopod dinosaur from the Upper Cretaceous of South Korea" (PDF). Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 259 (1): 1–24. doi:10.1127/0077-7749/2010/0102.
- Fearon, J.L.; Varricchio, D.J. (2015). "Morphometric analysis of the forelimb and pectoral girdle of the Cretaceous ornithopod dinosaur Oryctodromeus cubicularis and implications for digging". Journal of Vertebrate Paleontology. 35 (4): e936555. doi:10.1080/02724634.2014.936555.
- Huh, M.; Kim, J.G. (2010). "Koreanosaurus boseongensis". 한국고생물학회 정기총회 및 학술발표회 (in Korean): 21–26.
- Holtz, Thomas R. Jr. (2012). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages (PDF).
Winter 2011 Appendix
- ^ Horner, J. and Weishampel, D., 1988, "A comparative embryological study of two ornithischian dinosaurs", Nature (London), 332(No. 6161): 256-257 (1988)
- http://www.fossilworks.org/cgi-bin/bridge.pl?a=collectionSearch&taxon_no=67690&max_interval=Cretaceous&country=Canada&state=Alberta&is_real_user=1&basic=yes&type=view&match_subgenera=1
- Sullivan, R. M. (2003). "Revision of the dinosaur Stegoceras Lambe (Ornithischia, Pachycephalosauridae)". Journal of Vertebrate Paleontology. 23: 181–207. doi:10.1671/0272-4634(2003)23[181:rotdsl]2.0.co;2.
- Lambe L. M. (1902). New genera and species from the Belly River series (Mid-Cretaceous). Contributions to Canadian Paleontology, Geological Survey of Canada 3: 25–81.
- ^ Schott, R.K.; Evans, D.C. (2016). "Cranial variation and systematics of Foraminacephale brevis gen. nov. and the diversity of pachycephalosaurid dinosaurs (Ornithischia: Cerapoda) in the Belly River Group of Alberta, Canada". Zoological Journal of the Linnean Society. doi:10.1111/zoj.12465.
- ^ Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, New Jersey: Princeton University Press. pp. 268–269. ISBN 9780691167664.
- http://www.fossilworks.org/cgi-bin/bridge.pl?a=taxonInfo&taxon_no=38794
- Perle, A.; Maryańska, T.; Osmólska, H. (1982). "Goyocephale lattimorei gen. et sp. n., a new flat -headed pachycephalosaur (Ornithlschia , Dinosauria) from the Upper Cretaceous of Mongolia" (PDF). Acta Palaeontologica Polonica. 27 (1–4): 115–127.
- Mahito Watabe; Khishigjaw Tsogtbaatar; Robert M. Sullivan (2011). "A new pachycephalosaurid from the Baynshire Formation (Cenomanian-late Santonian), Gobi Desert, Mongolia" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 489–497.
- ^ Dyer, Aaron D.; Powers, Mark J.; Currie, Philip J. (2023). "Problematic putative pachycephalosaurids: Synchrotron µCT imaging shines new light on the anatomy and taxonomic validity of Gravitholus albertae from the Belly River Group (Campanian) of Alberta, Canada". Vertebrate Anatomy Morphology Palaeontology. 10 (1): 65–110. doi:10.18435/vamp29388. ISSN 2292-1389.
- Robert M. Sullivan (2003). "Revision of the dinosaur Stegoceras Lambe (Ornithischia, Pachycephalosauridae)". Journal of Vertebrate Paleontology. 23 (1): 181–207. doi:10.1671/0272-4634(2003)23[181:rotdsl]2.0.co;2.
- Maryańska, T.; Osmólska, H. (1974). "Pachycephalosauria, a new suborder of ornithischian dinosaurs" (PDF). Palaeontologica Polonica (30): 45−102.
- "Showcase Biological Specimen // MorphoSource".
- Goodwin, M. B.; Buchholtz, E. A.; Johnson, R. E. (1998). "Cranial anatomy and diagnosis of Stygimoloch spinifer (Ornithischia: Pachycephalosauria) with comments on cranial display structures in agonistic behavior". Journal of Vertebrate Paleontology. 18 (2): 363–375. doi:10.1080/02724634.1998.10011064.
- ^ Woodruff, D. Cary; Schott, Ryan K.; Evans, David C. (2023-11-15). "Two new species of small-bodied pachycephalosaurine (Dinosauria, Marginocephalia) from the uppermost Cretaceous of North America suggest hidden diversity in well-sampled formations". Papers in Palaeontology. 9 (6). e1535. doi:10.1002/spp2.1535. ISSN 2056-2799.
- Brown B.; Schlaikjer E. M. (1943). "A study of the troödont dinosaurs with a description of a new genus and four new species". Bulletin of the American Museum of Natural History. 82 (5): 115–149.
- Hatcher, J.B.; Lull, R.S.; Marsh, O.C.; Osborn, H. F. (1907). "The Ceratopsia". Monographs of the United States Geological Survey. XLIX. doi:10.5962/bhl.title.60500.
- ^ Jasinski, S. E.; Sullivan, R. M. (2011). "Re-evaluation of pachycephalosaurids from the Fruitland-Kirtland transition (Kirtlandian, late Campanian), San Juan Basin, New Mexico, with a description of a new species of Stegoceras and a reassessment of Texascephale langstoni" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 202–215.
- Robert M. Sullivan (2003) Revision of the dinosaur Stegoceras Lambe (Ornithischia, Pachycephalosauridae), Journal of Vertebrate Paleontology, 23:1, 181-207, DOI: 10.1671/0272-4634(2003)232.0.CO;2
- Sues, H. D. & Galton, P. M. (1987). "Anatomy and classification of the North American Pachycephalosauria (Dinosauria: Ornithischia)". Palaeontographica Abteilung A. 198: 1–40.
- Williamson, T. E.; Brusatte, S. L. (2016). "Pachycephalosaurs (Dinosauria: Ornithischia) from the Upper Cretaceous (upper Campanian) of New Mexico: A reassessment of Stegoceras novomexicanum". Cretaceous Research. 62: 29–43. doi:10.1016/j.cretres.2016.01.012.
- Fanti, F.; Cantelli, L.; Angelicola, L. (2018-04-01). "High-resolution maps of Khulsan and Nemegt localities (Nemegt Basin, southern Mongolia): Stratigraphic implications". Palaeogeography, Palaeoclimatology, Palaeoecology. THE LATE CRETACEOUS NEMEGT ECOSYSTEM: DIVERSITY, ECOLOGY, AND GEOLOGICAL SIGNATURE. 494: 14–28. Bibcode:2018PPP...494...14F. doi:10.1016/j.palaeo.2017.10.015. ISSN 0031-0182.
- Currie, P. J. (2016). Dinosaurs of the Gobi: Following in the footsteps of the Polish-Mongolian Expeditions. Palaeontologia Polonica, 67, 83-100.
- ^ Longrich, N.R.; Sankey, J. & Tanke, D. (2010). "Texacephale langstoni, a new genus of pachycephalosaurid (Dinosauria: Ornithischia) from the upper Campanian Aguja Formation, southern Texas, USA". Cretaceous Research. 31 (2): 274–284. doi:10.1016/j.cretres.2009.12.002.
- Head-ramming dino had ‘gears’ in skull
- Xu, X.; Tang, S.; Lin, S. (2016). "Paleostress inversion of fault-slip data from the Jurassic to Cretaceous Huangshan Basin and implications for the tectonic evolution of southeastern China". Journal of Geodynamics. 98: 31–52. doi:10.1016/j.jog.2016.03.013.
- ^ Hou, L., & Downs, T. B. W. (1977). A primitive pachycephalosaurid from the Cretaceous of Anhui, China, Wannanosaurus yansiensis gen. et sp. nov. Vertebrata PalAsiatica, 15(3), 1-9.
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Maryańska, Teresa (1990). "Pachycephalosauria". In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 564–577. ISBN 0-520-06727-4.
- "Abstract". Varricchio (2001); page 42.
- Kielan-Jaworowska (1970), "New Upper Cretaceous multituberculate genera from Bayn Dzak, Gobi Desert", Palaentologica Polonica 21, pp. 35-49.
- "Gobibaatar has page priority but was selected as the junior synonym by Kielan-Jaworowska in 1980," (McKenna & Bell, 1997)